

INTRODUCTION
The Neuquén Basin constitutes one of the most significant sedimentary basins in South America, with a well-documented stratigraphic succession spanning the Triassic through the Cenozoic (Kay & Ramos, 2006). During the Late Cretaceous, the tectonic and depositional dynamics of the basin were notably influenced by the ongoing Andean orogeny, resulting in the accumulation of fluvial and lacustrine sediments deposited within a context of progressive marine regression (Garrido, 2010). This phase is exemplified by the Neuquén Group, particularly the Río Colorado Subgroup, which marks the final continental sedimentation episode before the marine transgression of the Late Campanian (Legarreta & Gulisano, 1989). Among the units comprising the Río Colorado Subgroup, the Bajo de la Carpa Formation is of particular palaeontological interest. Dated to the Santonian (ca. 86–83 Ma), this formation is characterised by reddish, poorly cemented sandy and pelitic deposits, with colouration derived from iron oxide content (Garrido, 2011). The sedimentary facies reflect primarily continental depositional systems, indicative of fluvio-lacustrine environments with episodic subaerial exposure. The Neuquén Basin is renowned for its rich Mesozoic fossil record, providing critical insights to reconstruct the marine and continental palaeoenvironments of South America. The continental outcrops in its southern area yielded several vertebrate specimens of the Early Cretaceous, including flying reptiles, crocodiles, turtles, and dinosaurs (e.g., Gasparini et al., 2007; Calvo et al., 2004; Novas, 2009; Leanza et al., 2004), suggesting the presence of different palaeoecosystems with complex faunal compositions. The new sauropod specimens described in this contribution come from the deposits of the Bajo de la Carpa Formation. The first palaeontological reports from this unit date back to the late 19th and early 20th centuries, with the pioneering work of Roth (1898), and later systematic explorations by Bonaparte (1991), which laid the groundwork for subsequent discoveries. Over the past few decades, more comprehensive field campaigns have significantly expanded the known vertebrate assemblage of the formation, revealing a taxonomically diverse faunal community. This includes Titanosauria, Abelisauridae, Mesoeucrocodylia, and various non-dinosaurian vertebrates (Coria et al., 2013; Filippi et al., 2018). The titanosaurian sauropods, in particular, are well-represented, with several taxa described based on material from the Bajo de la Carpa Formation, including Bonitasaura salgadoi (Apesteguía, 2004), Rinconsaurus caudamirus (Calvo & González Riga, 2003), Overosaurus paradasorum (Coria et al., 2013), and Traukutitan eocaudata (Valieri & Calvo, 2011). Each of these taxa contributes to the growing understanding of titanosaur morphological diversity and biogeography within Gondwanan contexts. Nevertheless, several specimens remain indeterminate due to their fragmentary preservation, a common challenge in Cretaceous terrestrial vertebrate palaeontology (Cerda et al., 2012).
The Los Bastos locality, situated in the southeastern sector of the Neuquén Basin near Senillosa, represents a more recent addition to the roster of productive fossil sites in the region. First reported in 2017 by the Dirección Provincial del Patrimonio Cultural, the site initially yielded vertebrate material from the underlying Portezuelo Formation. Later studies expanded the known fossiliferous horizons to include the upper levels of the Bajo de la Carpa Formation and the base of the Anacleto Formation (Messina et al., 2023). These investigations have resulted in the recovery of both micro- and macro-vertebrate remains, with a particular abundance of titanosaurian elements collected from the sites LB1 and LB2 (Bellardini et al., 2021). The most recent investigations (Cori et al., 2023), started to establish a stratigraphic and palaeobiological framework for titanosaur remains from the Anacleto Formation within the Los Bastos area. However, titanosaurian material from the underlying Bajo de la Carpa Formation remains less thoroughly described and contextualised.
The present study aims to document and describe newly recovered titanosaurian material from the Bajo de la Carpa Formation at the Los Bastos locality (Neuquén Province, Argentina). This includes isolated and associated cranial and postcranial elements, housed in the collection of the Museo Municipal de Ciencias Naturales “Prof. Elizabeth Arocena” of Senillosa. By providing detailed osteological descriptions and taxonomic comparisons, this research seeks to expand the documented sauropod diversity from the Santonian of Patagonia. Through this approach, the study contributes not only to the taxonomy of South American titanosaurs but also to broader discussions of Gondwanan dinosaurian palaeobiology during the Late Cretaceous.
Institutional abbreviations
MAU, Museo Municipal Argentino Urquiza, Rincón de los Sauces, Neuquén, Argentina; MDPA, Museo del Desierto Patagónico de Añelo, Añelo, Neuquén, Argentina; MMCh, Museo Municipal “Ernesto Bachmann”, Villa El Chocón, Neuquén, Argentina; MMS, Museo Municipal de Ciencias Naturales “Prof. Elizabeth Arocena”, Senillosa, Neuquén, Argentina; MPCA, Museo Provincial “Carlos Ameghino”, Cipolletti, Río Negro, Argentina; MUC, Museo de Ciencias Naturales de la Universidad Nacional del Comahue, Neuquén, Argentina; PV, Paleovertebrate collection; MMS-PV, acronym for vertebrate palaeontology specimens housed at the MMS.
GEOLOGICAL SETTING
The fossil material analysed in this study originates from the Bajo de la Carpa Formation, a Santonian-aged unit situated at the base of the Río Colorado Subgroup within the Neuquén Group. The Neuquén Group, a prominent stratigraphic succession in the Neuquén Basin of west-central Argentina (Fig. 1A), preserves a rich record of terrestrial and marginal marine ecosystems from the Late Cretaceous and is subdivided into three subgroups: the Río Limay, Río Neuquén, and Río Colorado (Legarreta & Gulisano, 1989; Garrido, 2010). The Bajo de la Carpa Formation was initially considered a member of a broader Río Colorado Formation (Cazau & Uliana, 1973), which also encompassed the Anacleto and Allen formations. However, subsequent revisions by Ramos (1981) elevated the Bajo de la Carpa and Anacleto units to formational rank within the Río Colorado Subgroup, separating the overlying Allen Formation and reassigning it to the younger Malargüe Group. As presently defined, the Río Colorado Subgroup comprises the Bajo de la Carpa and Anacleto formations and marks the final continental phase of sedimentation before the Late Campanian marine ingression that characterises the transition to the Malargüe Group (Ramos & Folguera, 2005). Lithologically, the Bajo de la Carpa Formation is dominated by a series of weakly cemented psammitic and pelitic deposits. These are composed primarily of fine to medium quartzitic sandstones interbedded with reddish to yellowish mudstones and siltstones, commonly interpreted as the products of fluvial and ephemeral lacustrine environments (Garrido, 2010; Leanza et al., 2004). The iron oxide content is responsible for the distinctive reddish colouration seen in many exposures, particularly toward the centre of the basin, where oxidising conditions likely prevailed.
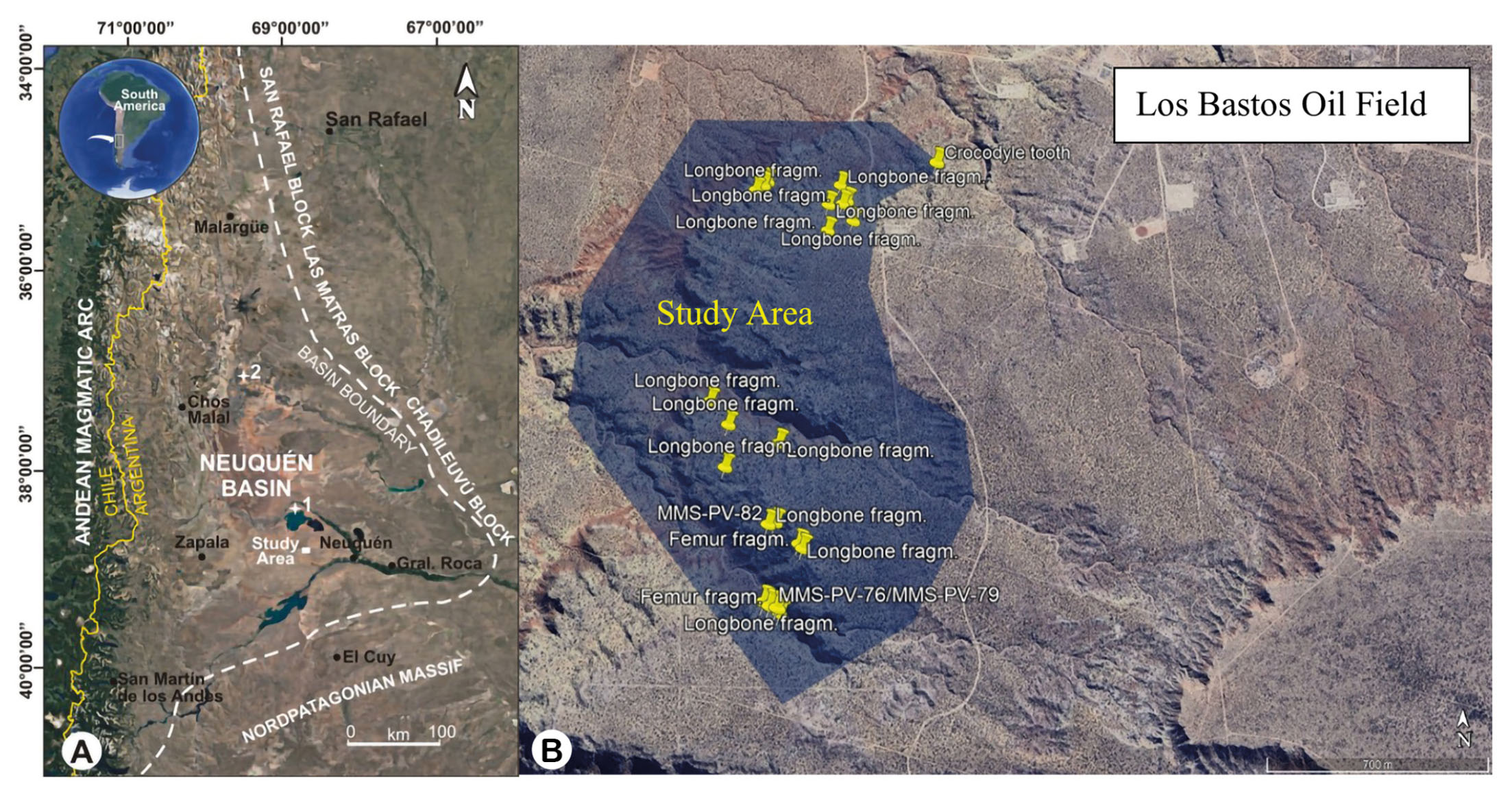
- (A) On the left, extension of the Neuquén Basin. Modified from Bellardini et al. (2021). (B) On the right, polygonal representation of the prospected area of the Los Bastos locality with the geographical coordinates of both collected specimens and other findings.
The formation displays a characteristic vertical succession of facies, with sandstone bodies representing channel-fill and sheet flood deposits, and finer-grained units interpreted as overbank and floodplain deposits (Garrido, 2011). These depositional settings are consistent with seasonal semi-arid conditions that dominated the Neuquén Basin during the Santonian (Legarreta & Gulisano, 1989). Sedimentary structures such as crossbedding, mud cracks, and root traces indicate periodic exposure and soil formation in the floodplain environments. Stratigraphically, the Bajo de la Carpa Formation conformably overlies the Portezuelo Formation, which belongs to the Río Neuquén Subgroup. It is, in turn, conformably overlain by the Anacleto Formation. The total thickness of the Bajo de la Carpa Formation varies across the basin but generally ranges between 60 and 150 meters (Garrido, 2010). Exposures of the unit are particularly well-developed along the eastern margins of the Neuquén Basin, where it caps the ‘Bardas’, steep escarpments that mark the physiographic transition between different sedimentary packages.
The fossil specimens described in this study were collected from surface exposures of the Bajo de la Carpa Formation at the Los Bastos locality (Fig. 1B). These sediments are typically poorly consolidated and weakly cemented, which has contributed to the high degree of weathering observed in the recovered bones. The fragile condition of the material is consistent with post-depositional exposure and erosion under arid to semi-arid conditions, which can lead to the fragmentation and deflation of fossiliferous horizons (Bellardini et al., 2021). Nonetheless, these exposures have proven productive, yielding both macrofossil and microfossil vertebrate remains, and offer significant potential for future discoveries within the Santonian terrestrial ecosystems of northern Patagonia.
The geological and taphonomic context of the Bajo de la Carpa Formation provides essential information for interpreting the palaeoenvironmental conditions under which the Los Bastos titanosaurians were preserved. The association of titanosaurian remains with overbank mudstones and fluvial sandstones supports the interpretation that these animals inhabited dynamic floodplain environments characterised by intermittent fluvial activity and seasonal climatic variability. Moreover, the conformable nature of the contact with the overlying Anacleto Formation ensures that stratigraphic correlations between titanosaur-bearing horizons can be established with relative precision across the basin.
MATERIALS AND METHODS
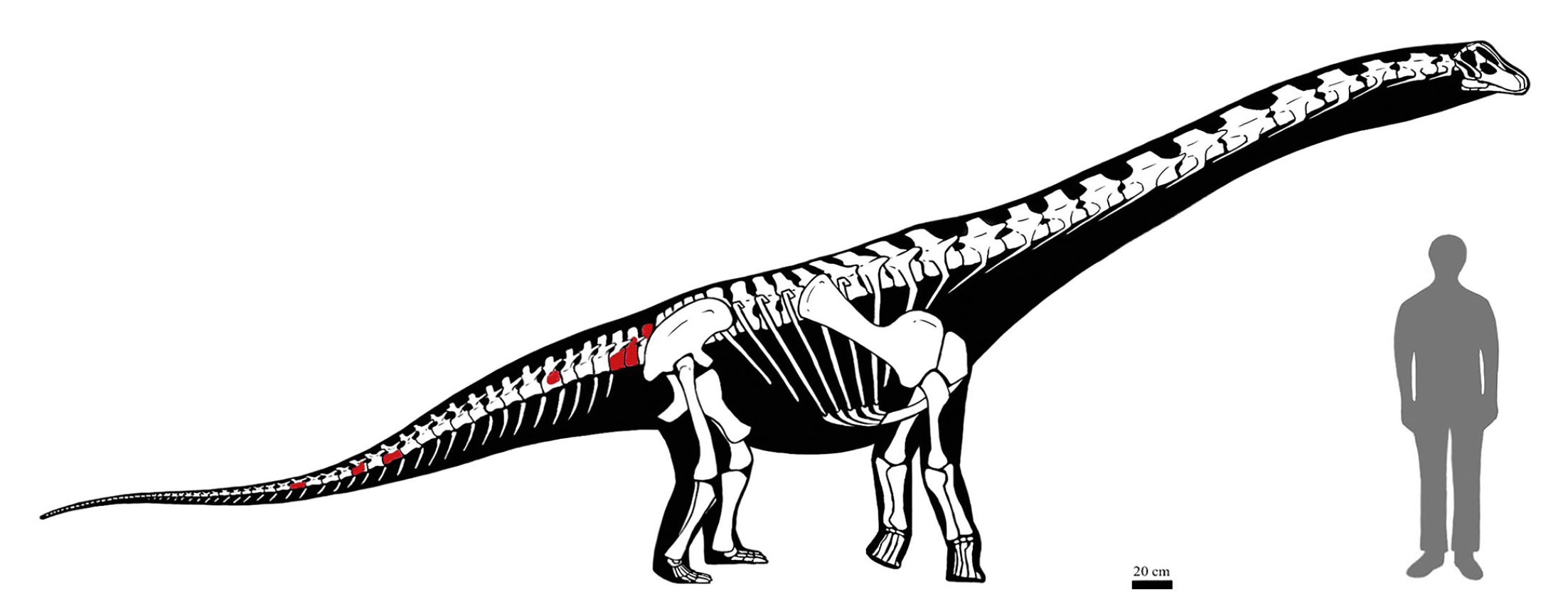
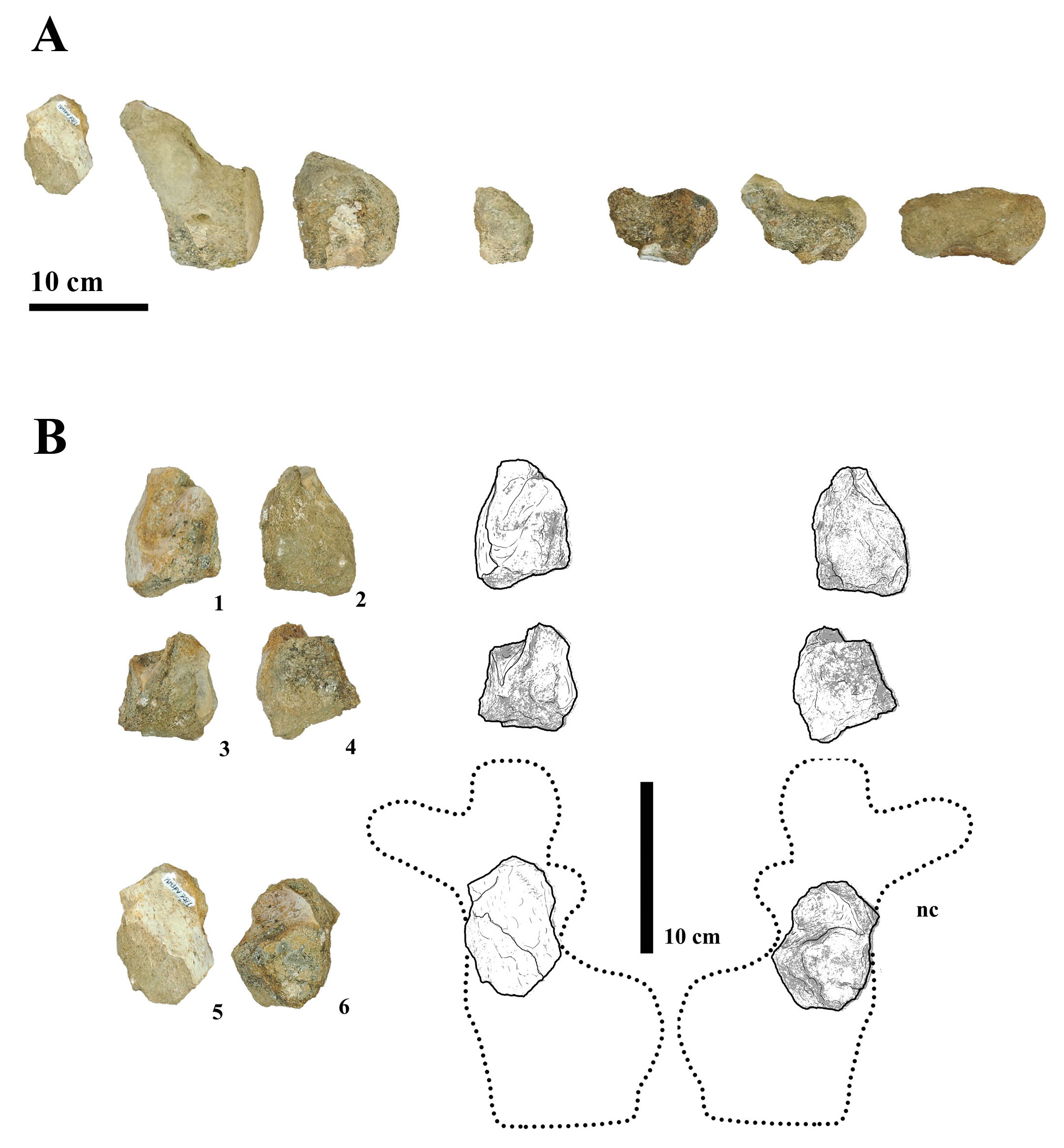
The titanosaurian fossil material described in this study consists of both cranial and postcranial elements (10 elements in total) recovered from the Los Bastos locality, situated near the town of Senillosa in the Neuquén Province of Argentina (see Tab. 1). The fossils originate from fieldwork conducted during two exploratory campaigns targeting the Las Chivas locality, which occupies the northern portion of the Los Bastos area. The fieldwork were designed to investigate previously undocumented exposures of fossiliferous sediments and to recover new specimens contributing to the study of titanosaurian diversity in the region. The fossil elements include a series of seven associated caudal vertebrae attributed to a single individual MMS-PV-75 (Fig. 2), and three elements of the pelvic girdle (catalogued as MMS-PV-82), in addition to several isolated vertebral, cranial, and limb fragments. These remains were collected during recent systematic surveys, except for the initial four specimens listed, which were recovered in earlier campaigns. Each finding was thoroughly documented with high-resolution photographs and georeferenced using GPS coordinates. Linear measurements were taken from the better-preserved specimens MMS-PV-39, MMS-PV-75 (elements 2, 3, 5, 6, and 7), and MMS-PV-76 to calculate both vertical and horizontal elongation indices, following the methods of Wilson & Sereno (1998) and Upchurch (1998), respectively (see Tab. 2). These measurements consist in the height and width of the posterior articular surface, as well as the anteroposterior length of the centrum. However, due to erosion or absence of key portions of the vertebral surfaces, the measurements had to be approximated and therefore could not provide reliable data for comparative purposes. All recovered specimens are housed in the Museo Municipal de Ciencias Naturales “Prof. Elizabeth Arocena” of Senillosa and are labelled using the institutional prefix MMS (Museo Municipal de Senillosa), followed by the collection category acronym PV (Vertebrate Palaeontology) and an individual inventory number. Each bony element included in this study was examined in detail to assess its anatomical features and potential taxonomic affinities. This process involved direct comparisons with previously described material from the Bajo de la Carpa Formation and other stratigraphically equivalent or adjacent units within the Neuquén Group. Specimens were compared with material held in a variety of regional collections, including the Museo Municipal Ernesto Bachmann (MMCh), Villa El Chocón; the Museo Provincial Carlos Ameghino (MPCA), Cipolletti; the Museo Municipal Argentino Urquiza (MAU), Rincón de los Sauces; the Museo del Desierto Patagónico de Añelo (MDPA), Añelo; and the Museo de Ciencias Naturales de la Universidad Nacional del Comahue (MUC), located in Neuquén Capital. The comparative sample included both cranial and postcranial elements from Titanosauria, particularly those taxa previously recovered from the Bajo de la Carpa and Anacleto formations, as well as elements belonging to other neosauropod groups such as Rebbachisauridae, which were consulted to assess broader morphological patterns. Anatomical terminology for vertebral and appendicular elements follows standard sources used in sauropod palaeontology, including Romer (1956), Wilson & Sereno (1998), Harris (2004), Upchurch et al. (2004), and Wilson (2006). Special attention was given to vertebral proportions, pneumaticity, lamination patterns, and articular surface morphology, which are critical for establishing serial and positional identity within the axial skeleton.
| Specimens | Material | Taxonomic attribution | Geographical reference |
|---|---|---|---|
| MMS-PV-39 | Caudal vertebra | Titanosauria indet. | 38°51’30,8”S/68°39’48,1”W |
| MMS-PV-40 | Caudal vertebra | Titanosauria indet. | 38°51’31,4”S/68°39’49,8”W |
| MMS-PV-41 | Caudal vertebra | Titanosauria indet. | 38°51’30”S/68°39’38”W |
| MMS-PV-75/01-07 | Seven caudal vertebrae | Titanosauria indet. | 38°52’19,17”S/68°39’52,49”W |
| MMS-PV-76 | Caudal vertebra | Titanosauria indet. | 38°52’18,5”S/68°39’53,9”W |
| MMS-PV-77 | Right frontal | Titanosauria indet. | 38°51’27.8”S/68°39’48”W |
| MMS-PV-78 | Caudal neural spine | Titanosauria indet. | 38°51’29,5”S/68°39’55,9”W |
| MMS-PV-79 | Right metacarpal IV | Titanosauria indet. | 38°52’18,5”S/68°39’53,9”W |
| MMS-PV-80 | Right ulna diaphysis | Titanosauria indet. | 38°51’28”S/68°39’55,3”W |
| MMS-PV-82/01-03 | pelvic girdle elements | Titanosauria indet. | 38°52’09,5”S/68°39’53,3”W |
| Specimen | Cotyle height | Cotyle width | Centrum length | Vertical elongation index | Horizontal elongation index |
|---|---|---|---|---|---|
| MMSPV39 | 7 | 8.5 | 9.5 | 1.36 | 1.12 |
| MMSPV75-2 | 8 | 8.5 | 8 | 1 | 0.94 |
| MMSPV75-3 | 7 | 8 | 8.5 | 1.21 | 1.06 |
| MMSPV75-5 | 6.5 | 7 | 9.5 | 1.46 | 1.36 |
| MMSPV75-6 | 6.5 | 8.5 | 10.5 | 1.61 | 1.23 |
| MMSPV75-7 | 6.5 | 8.5 | 13.5 | 2.07 | 1.59 |
| MMSPV76 | 5.5 | 6.5 | 10.5 | 1.9 | 1.61 |
- Reconstruction of specimen MMS-PV-75 alongside a 185 cm tall human figure, based on the skeletal elements recovered and described in this study (highlighted in red), and supplemented through comparative analysis with morphologically similar titanosaurian clades, including Rinconsauria and Aeolosaurines. Paleoart by Riccardo Serena de Maria.
DESCRIPTION AND COMPARISON
Systematic Palaeontology
Macronaria Wilson & Sereno, 1998.
Titanosauria Bonaparte & Coria, 1993.
All the specimens described in the present study have been assigned to Titanosauria indet.
MMS-PV-39
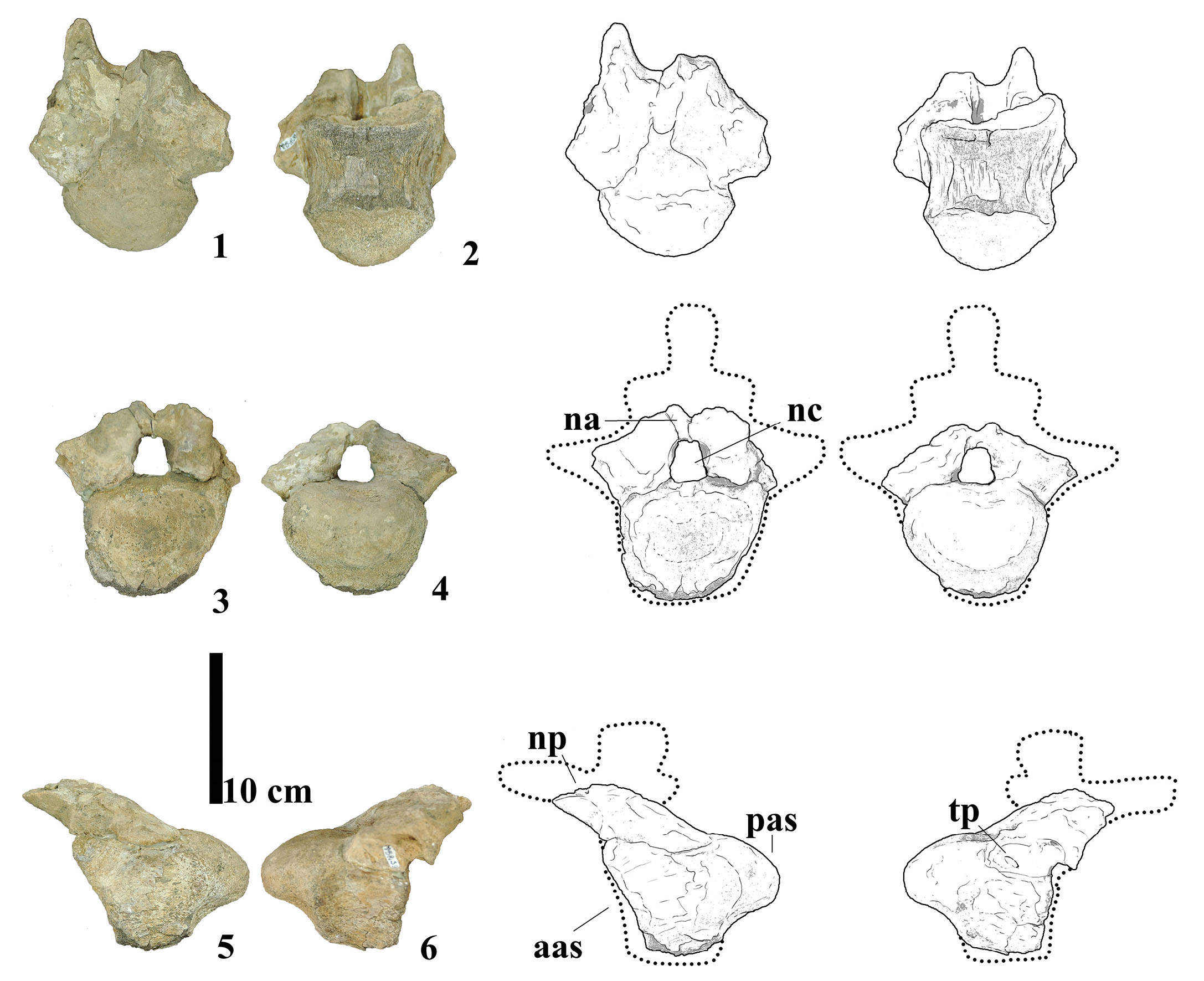
This specimen comprises a partially preserved anterior caudal vertebra (Fig. 3). The centrum is largely intact, although the anteroventral region is missing, and it preserves parts of the transverse processes, both neural pedicels, and the proximal bases of the prezygapophyses. The centrum exhibits a strong procoelous condition, with a concave anterior articular face and a posterodorsally convex posterior condyle. In anterior view, the centrum is wider than tall, similarly to the condition seen in the Aeolosaurine Panamericansaurus schroederi (Calvo & Porfiri, 2010). In lateral view, the anterior face is steeply inclined anteriorly. The anteroventral inclination of the anterior articular surface is commonly found across members of the clades Rinconsauria and Aeolosaurini in literature. This condition is more evident in Gondwanatitan faustoi (Kellner & de Azevedo, 1999), Aeolosaurus rionegrinus (Powell, 1987), and Punatitan coughlini (Hechenleitner et al., 2020), and is less marked in MMS-PV-39 similarly to Rinconsaurus caudamirus (Calvo & González Riga, 2003), Overosaurus paradasorum (Coria et al., 2013), Trigonosaurus pricei (Campos et al., 2005), Uberabatitan ribeiroi (Salgado & Carvalho, 2008), Arrudatitan maximus (Santucci & de Arruda-Campos, 2011), and Adamantisaurus mezzalirai (Santucci & Bertini, 2006). Outside of these two groups, it is also present in Bonitasaura salgadoi (Apesteguía, 2004), Rapetosaurus krausei (Curry Rogers & Forster, 2001), and Narambuenatitan palomoi (Filippi et al., 2011). The lateral surfaces of the centrum lack any evidence of pneumatic features such as fossae or foramina, presenting instead a mildly convex dorsoventral profile and slight anteroposterior concavity. Ventrally, the centrum is transversely convex and narrow, with no indication of chevron facets (haemapophyses), a midline keel, or ventrolateral ridges. The absence of these features is notable, as ventrolateral ridges are present in some derived titanosaurs such as Trigonosaurus pricei (Campos et al., 2005) but are absent in others like Gondwanatitan faustoi (Kellner & Azevedo, 1999) and Bonitasaura salgadoi (Apesteguía, 2004). The neural arch is positioned on the anterior half of the centrum and is inclined anterodorsally. The preserved neural canal is elliptical in shape and mediolaterally compressed. Diapophyseal laminae are not discernible due to damage, and the transverse processes appear triangular in dorsal view, with the preserved base projecting posterodorsally. The pedicels are low and robust, and neither the centrum nor the arch shows signs of internal pneumaticity (i.e., camellae or camerae), a feature consistent with basal titanosaurian conditions (Wilson, 2006). Comparative analysis indicates that the observed features are morphologically consistent with an anterior caudal position for this vertebra. The combination of a short centrum, an anteriorly placed neural arch, and the lack of haemapophyses is consistent with proximal caudals, probably amongst the first 3 to 4 vertebrae, both in South American colossosaurians such as Uberabatitan ribeiroi (Salgado & De Souza Carvalho, 2008) and Patagotitan mayorum (Carballido et al., 2017) and in European titanosauriformes such as Lusotitan atalaiensis (Mannion et al., 2013) and Lirainosaurus astibiae (Díez Díaz et al., 2013). The transversely convex ventral surface aligns more closely with Rinconsaurus caudamirus (Calvo et al., 2003) than with the flat or concave ventral surfaces seen in Overosaurus paradasorum (Coria et al., 2013) or Aeolosaurus rionegrinus (Powell, 2003). The absence of pneumatic structures and the solid texture of the centrum further distinguish this specimen from saltasaurines such as Saltasaurus loricatus (Bonaparte & Powell, 1980), in which internal pneumaticity is often well-developed.
- Photographs and line drawings of caudal vertebra MMS-PV-39 in (1) dorsal, (2) ventral, (3) anterior, (4) posterior, (5) left and (6) right views. Abbreviations: aas, anterior articular surface; na, neural arch; nc, neural canal; np, neural pedicels; pas, posterior articular surface; tp, transverse process.
MMS-PV-40
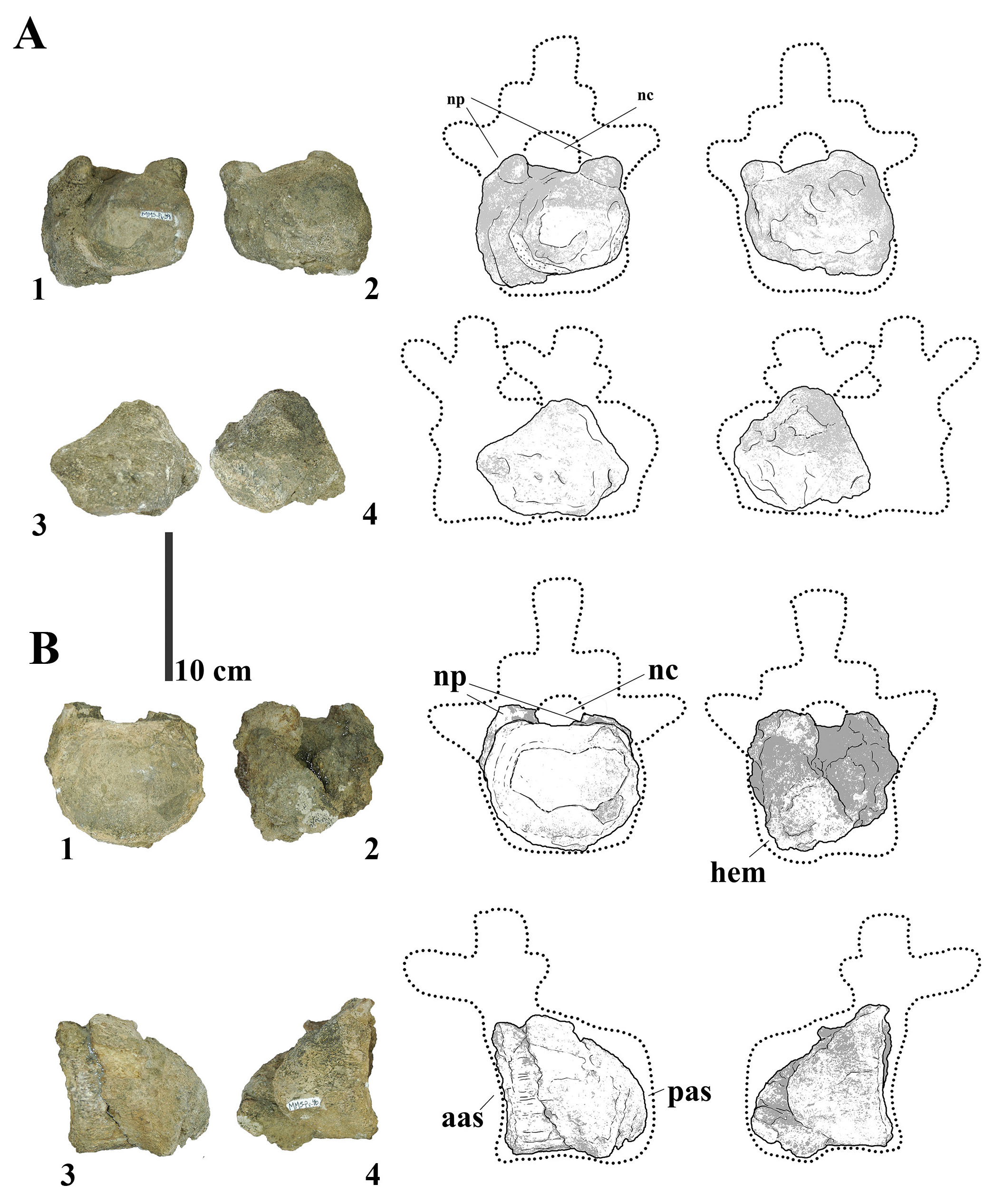
MMS-PV-40 consists of two fused caudal vertebral centra, preserved in a highly fragmented and diagenetically deformed state (Fig. 4A). The distalmost vertebra retains the basal portions of the neural pedicles, which are positioned widely apart, resulting in a notably broad neural canal. No camellate internal structure is visible in the broken sections of the centrum walls; instead, the bone matrix appears compact and solid. A ventral projection interpreted as a haemapophysis is preserved on the distalmost centra. The presence of fused caudal vertebrae is an uncommon but documented condition among titanosaurians and may result from either pathological processes or ontogenetic fusion during skeletal maturation (Cerda et al., 2015; García et al., 2019). Notably, similar fusion has been described in Epachthosaurus sciuttoi (Martínez et al., 2004), among other basal titanosauriforms.
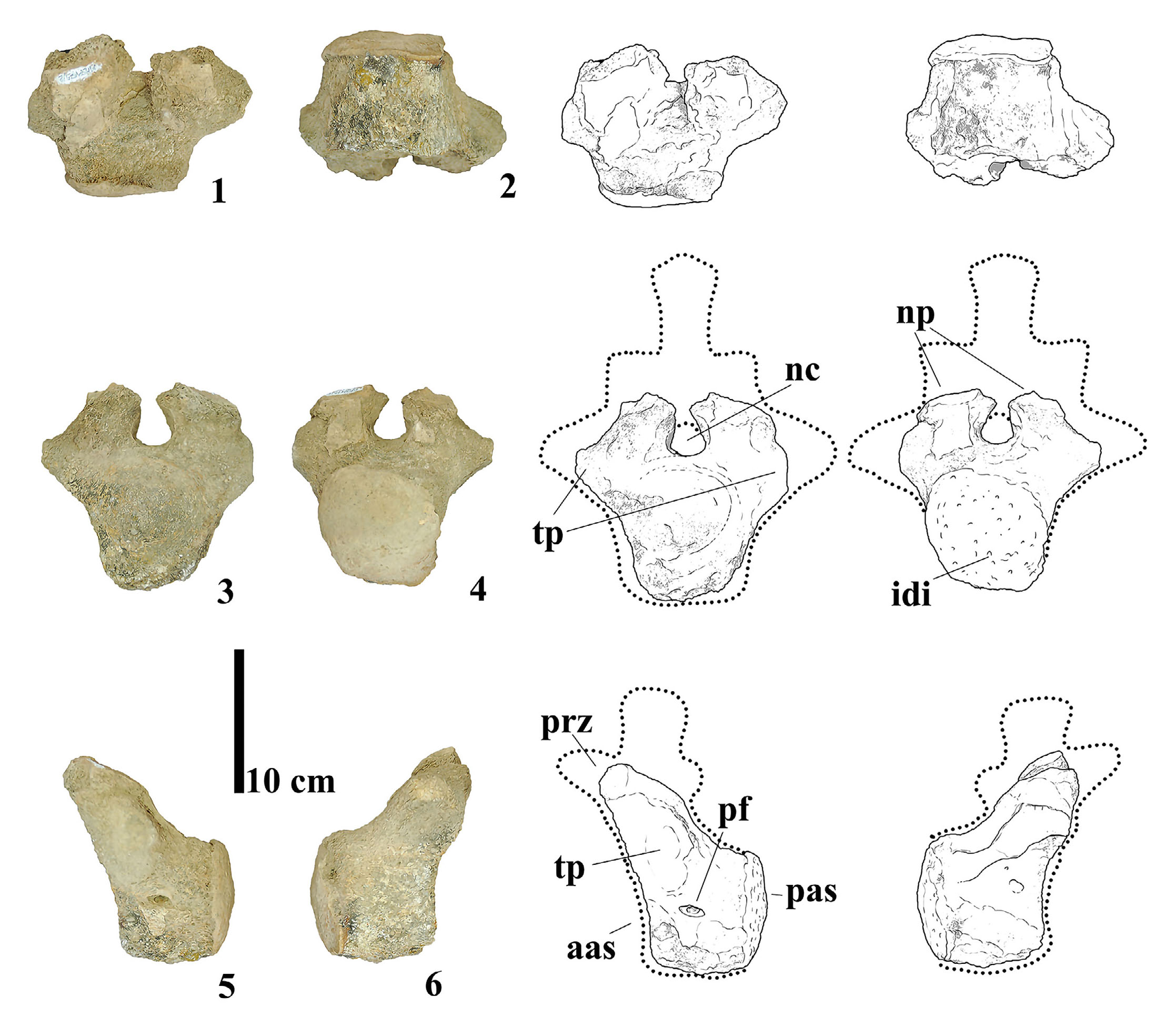
- (A) Photographs and line drawings of the fused caudal vertebrae of MMS-PV-40 in (1) anterior, (2) posterior, (3) left and (4) right views. (B) Photographs and line drawings of caudal vertebra MMS-PV-41 in (1) anterior, (2) posterior, (3) left and (4) right views. Abbreviations: aas, anterior articular surface; hem, haemapophysis; nc, neural canal; np, neural pedicels; pas, posterior articular surface.
MMS-PV-41
This specimen includes the anterior articular face of a caudal centrum and the base of the neural pedicels (Fig. 4B), it appears to be procoelic due to the shape of the eroded portion. The articular surface is transversely broad and dorsoventrally compressed, with a deeply concave dorsal portion. A protuberance on the right ventral margin, interpreted as an eroded haemapophysis, adds further evidence for a caudal vertebral identification. Like MMS-PV-39 and MMS-PV-40, the internal structure of the centrum is compact and shows no signs of internal pneumaticity. The overall morphology of the articular face is consistent with anterior to middle caudal vertebrae of Bonitasaura salgadoi (Apesteguía, 2004), particularly in the combination of width, concavity, and dorsoventral asymmetry. The lateral placement of the haemapophyses is more similar to Rinconsaurus caudamirus (Calvo et al., 2003) than to Overosaurus paradasorum, which tends to exhibit them more medially positioned.
MMS-PV-75
This specimen consists of a sequence of seven associated but badly preserved caudal vertebrae, recovered in association from the Bajo de la Carpa Formation at the Los Bastos locality (Fig. 5A). Although the neural arches are largely eroded or missing, preservation of the centra, transverse processes, and portions of the neural pedicels allows for partial morphological assessment. The vertebrae vary from anterior to mid-caudal in position and show a progression of characters useful for positional inference and taxonomic evaluation.
(A) - Caudal series of MMS-PV-75 in left view, in order from proximalmost to distalmost (left to right). (B) Photographs and line drawings of neural pedicel MMS-PV-75/01 in (1) dorsal, (2) ventral, (3) anterior, (4) posterior, (5) left and (6) right views. Abbreviations: nc, neural canal.
MMS-PV-75/01 - Represented by the left neural pedicel of a caudal vertebra, this element is preserved in isolation but is believed to occupy one of the most anterior positions within the sequence, potentially the first or second caudal (Fig. 5B). The robust and high pedicel is consistent with the morphology observed in anterior caudal series of titanosaurians such as Overosaurus paradasorum and Rinconsaurus caudamirus, as well as Aeolosaurus rionegrinus (Powell, 2003) and Gondwanatitan faustoi (Kellner & Azevedo, 1999), as reported in the literature.
MMS-PV-75/02 - This vertebra preserves part of the centrum, including the posterior articular surface, the proximal portions of the transverse processes, and fragments of the neural arch (Fig. 6). The centrum is as long as tall in lateral view. The anterior articular face is concave and oval, while the posterior face is sub-rectangular and flat, a condition referred to in the literature as procoelous-opisthoplatyan (Tidwell et al., 2001). This morphology is observed in basal titanosauriforms and contrasts with the strongly procoelous caudals of more derived titanosaurs (e.g., Saltasaurus, Neuquensaurus). Small pneumatic foramina (pneumatopores) are present near the neurocentral suture on both sides, a feature commonly observed in anterior caudals of Rinconsaurus caudamirus (Calvo et al., 2003) and Rocasaurus muniozi (Salgado & Azpilicueta, 2000). The transverse processes are ventrally expanded and display a characteristic bulge near their base, a feature known in Uberabatitan ribeiroi (Salgado & De Souza Carvalho, 2008). The ventral surface of the centrum is flat in lateral view but transversely convex anteriorly and lacks ridges or chevron facets.
- Photographs and line drawings of caudal vertebra MMS-PV-75/02 in (1) dorsal, (2) ventral, (3) anterior, (4) posterior, (5) left, and (6) right views. Abbreviations: aas, anterior articular surface; idi, intervertebral disks impressions; nc, neural canal; np, neural pedicels; pas, posterior articular surface; pf, pneumatic foramina; prz, prezygapophysis; tp, transverse processes.
MMS-PV-75/03 - This element preserves a portion of the centrum and the basal portion of the right transverse process (Fig. 7). In lateral view, the centrum is only slightly longer than tall. On both lateral sides, circular pneumatopores are visible. Vascular foramina are observed on the ventral surface. A ventral projection, interpreted as a haemapophysis, is preserved posteriorly. The combination of short anteroposterior length, pneumatic features, and the presence of haemapophyses indicate that this represents an anterior caudal vertebra, likely the third of the preserved sequence.
- Photographs and line drawings of caudal centrum MMS-PV-75/03 in (1) dorsal, (2) ventral, (3) anterior, (4) posterior, (5) left, and (6) right views. Abbreviations: aas, anterior articular surface; hem, haemapophysis; nc, neural canal; paf, posterior articular surface; pf, pneumatic foramina; tp, transverse process; vf, vascular foramina.
MMS-PV-75/04 - This fragmentary vertebra preserves part of the centrum and neural pedicels, with limited cortical surface remaining (Fig. 8A). The anterior articular surface is taller than wide, and although most morphological details are eroded, the centrum’s proportions suggest an intermediate position, likely among the anteriormost middle caudals.
- (A) Photographs and line drawings of caudal vertebra MMS-PV-75/04 in (1) anterior, (2) posterior. (B) Photographs and line drawings of caudal vertebra MMS-PV-75/05 in (1) left and (2) right views. (C) Photographs and line drawings of caudal vertebra MMS-PV-75/06 in (1) left and (2) right views. (D) Photographs and line drawings of caudal centrum MMS-PV-75/07 in (1) left and (2) right views. Abbreviations: aas, anterior articular surface; hem, haemapophysis; nc, neural canal; np, neural pedicels; pas, posterior articular surface; tp, transverse process.
MMS-PV-75/05 - This procoelic vertebra retains the posterior portion of the centrum and the neural pedicels (Fig. 8B). The preserved condyle is dorsally oriented, with its apex extending above the dorsal centrum margin in lateral view. The left posterior haemapophysis is partially preserved and placed laterally. All these features are indicative of a middle caudal vertebra and resemble the condition found in Gondwanatitan and Aeolosaurus, especially the vertical orientation of the condyle and laterally positioned peduncles.
MMS-PV-75/06 - This procoelic vertebra preserves most of the centrum and portions of the neural (Fig. 8C). The centrum is longer than tall. In dorsal view, the posterior condyle bears a shallow dorsal notch. Vascular foramina are present on both lateral and ventral surfaces. The neural arch is anteriorly located and preserves low, curved pedicels. All of the characters above support a distal position in the middle caudal region of this element.
MMS-PV-75/07 - The distalmost vertebra of the series, MMS-PV-75/7 includes a deformed centrum and part of the left transverse process (Fig. 8D). The procoelic centrum is wider than tall and rectangular in anterior view, with a relatively anteroposteriorly elongated profile in lateral view. The transverse process is robust and positioned lower on the lateral surface of the vertebra than in other elements of the series. A partially preserved haemapophysis is observed ventrally. The forementioned proportions and transverse process development indicate that this element represents a distal segment of the middle caudal region.
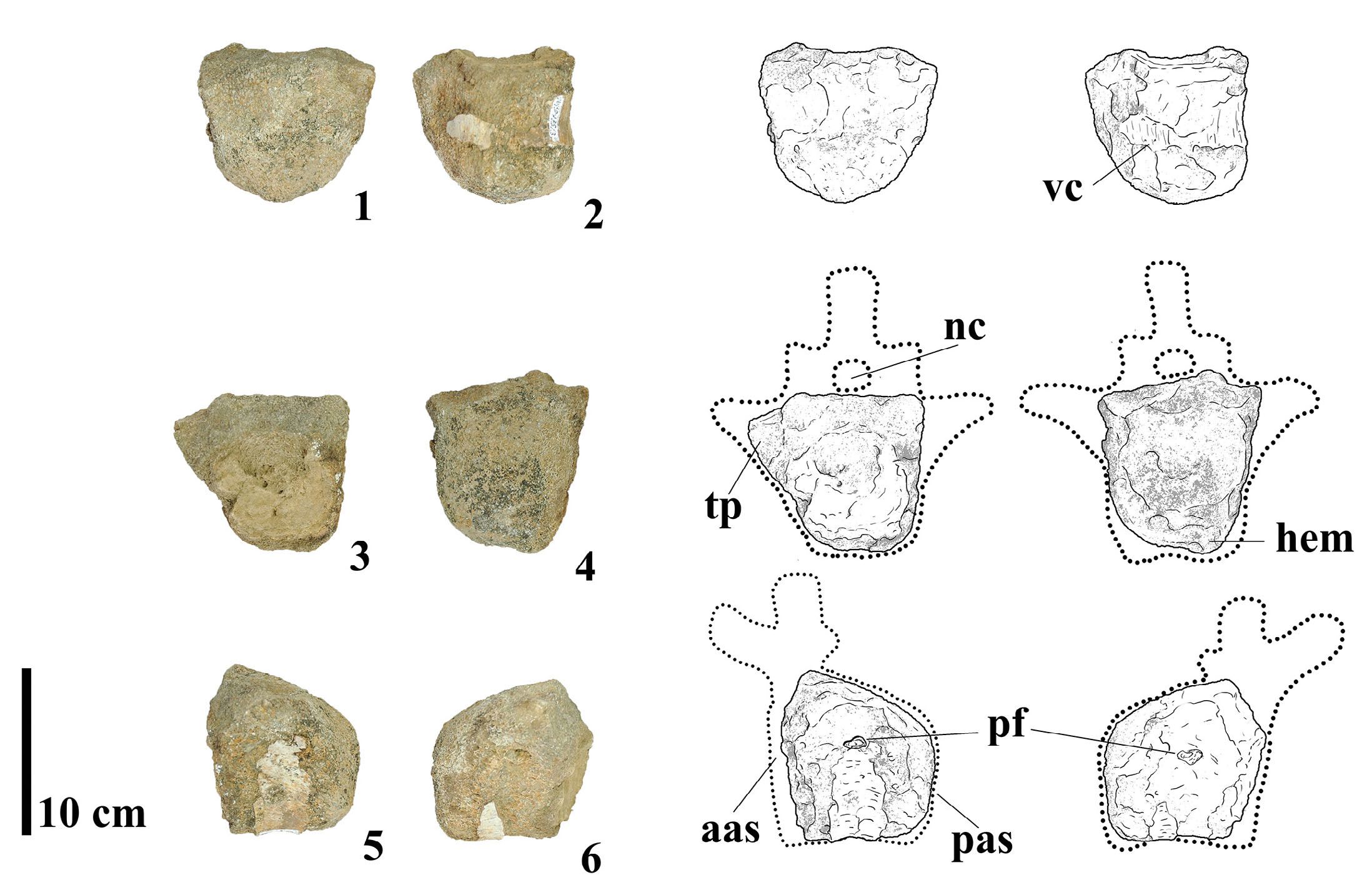
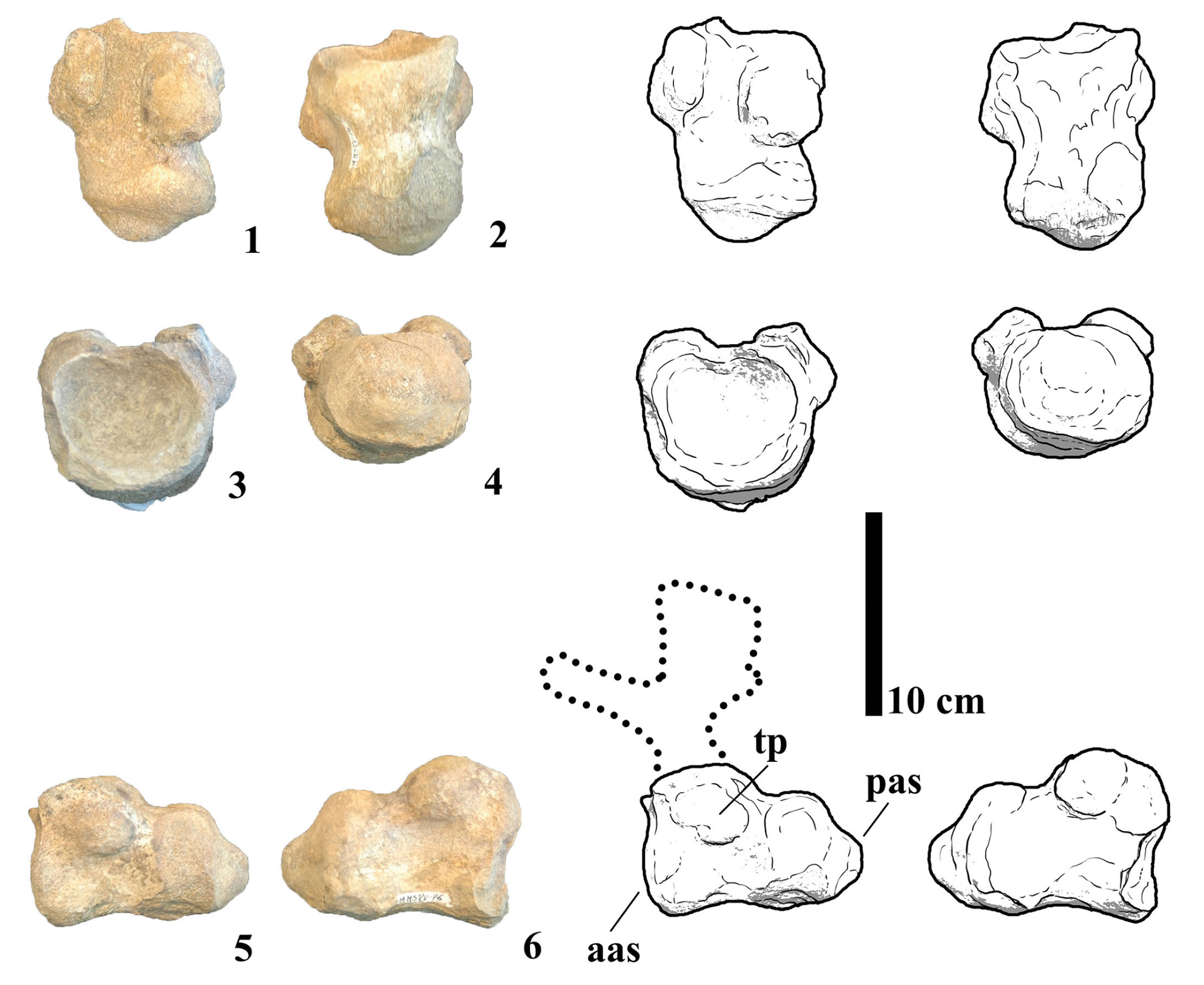
MMS-PV-76
This specimen comprises a relatively complete caudal vertebra, preserving most of the centrum, both transverse processes, and the basal portions of the neural pedicels (Fig. 9). In lateral view, the centrum is longer than tall, indicating a likely anterior to middle caudal position. The anterior articular surface is rounded and deeply concave, while the posterior surface is convex, with particular emphasis on the ventral half in lateral view. The lateral sides of the centrum are dorsoventrally convex and slightly concave anteroposteriorly. The ventral margin is gently concave in profile and transversely convex, lacking haemapophyses, a ventral keel, or longitudinal ridges. Notably, no external pneumatic features, such as lateral fossae or foramina, are present. The transverse processes are laterally projecting, flat, and situated low on the lateral surfaces of the centrum rather than elevated on the neural arch. The neural pedicels are slender, low, without reaching the neural arch height, a feature characteristic of anterior to middle caudal vertebrae among titanosaurs (Wilson, 2006). Comparative analysis suggests that MMS-PV-76 closely resembles the middle caudal vertebrae of Narambuenatitan palomoi (Filippi et al., 2011), due to the absence of pneumatic features, and the shape of the transverse processes. However, the specimen differs from rinconsaurians such as Gondwanatitan faustoi and Aeolosaurus rionegrinus in having a straighter anterior articular face, as rinconsaurians often exhibit a pronounced anterodorsal inclination in this region (Santucci & Arruda-Campos, 2011).
- Photographs and line drawings of caudal vertebra MMS-PV-76 in (1) dorsal, (2) ventral, (3) anterior, (4) posterior, (5) left and (6) right views. Abbreviations: aas, anterior articular surface; np, neural pedicels; pas, posterior articular surface; tp, transverse process.
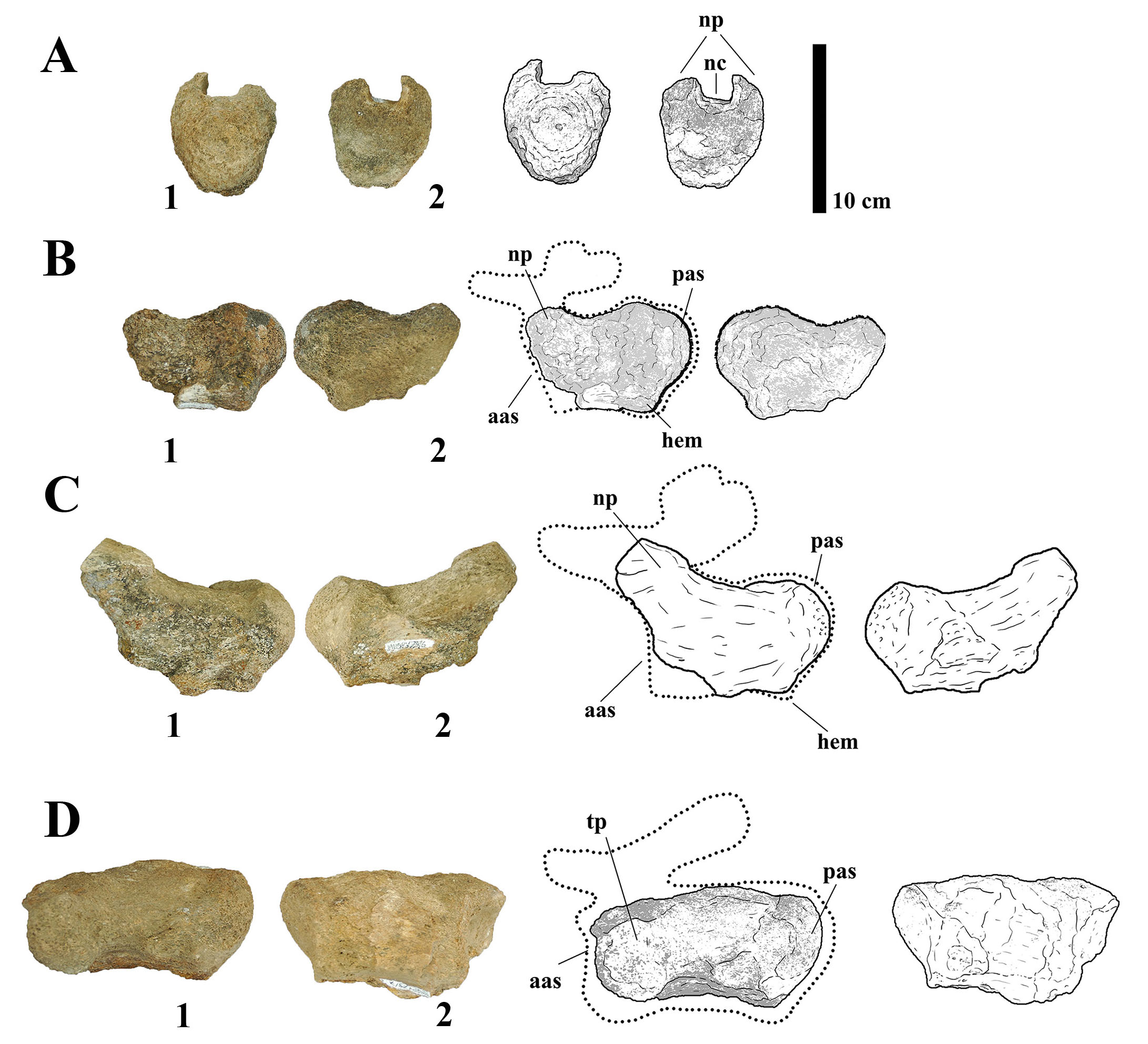
MMS-PV-77
This specimen consists of a nearly complete right frontal, preserving the dorsal surface, orbital roof, and a portion of the endocranial cavity (Fig. 10A). In anterior view, the frontal is dorsomedially convex, forming a dome that spans approximately the medial two-fifths of the element. The orbital roof is gently arched in lateral profile and transversely concave, flaring laterally to meet the contact surface with the prefrontal and nasal. A deep lateral notch is present on the anterior margin, interpreted as the articulation for the prefrontal, whereas a faint transverse facet medial to the notch likely marks the contact area for the nasal. The bone displays typical titanosaurian features, including the dorsomedial convexity observed in Saltasaurus loricatus (Bonaparte & Powell, 1980) and the wide orbital roof morphology present in Rapetosaurus krausei (Curry Rogers & Forster, 2004). The frontal is approximately as anterodorsally long as transversely wide, differently from the one described for the Australian titanosaur Diamantinasaurus matildae (Poropat et al., 2023) which is wider than long. The orbital margin is ornamented as in Bonitasaura, Rapetosaurus and Nemegtosaurus, unlike the basal Lithostrotian Sarmientosaurus mustacchioi (Martinez et al., 2016), which has a smooth margin. The depth and orientation of the prefrontal notch are less pronounced than in Nemegtosaurus mongoliensis (Nowinski, 1971), which shows a more deeply inset articulation, but more prominent than in Camarasaurus species, where the frontal margins are smoother and more rounded.
- (A) Photographs and line drawings of left frontal MMS-PV-77 in (1) dorsal and (2) ventral views. Abbreviations: cc, cranial cavity; ls/obs, laterosphenoid/orbitosphenoid suture; n, nasal facet; p, parietal facet; pfr, prefrontal facet. (B) Photographs and line drawings of neural spine MMS-PV-78 in (1) left and (2) right views. Abbreviations: poz, postzygapophyses. (C) Photographs and line drawings of IV right metacarpal MMS-PV-79 in (1) proximal and (2) distal views. Abbreviations: am, anterior margin; lm, lateral margin. (D) Photographs and line drawings of ulna MMS-PV-80 in (1) medial and (2) lateral views. Abbreviations: raa, radial articulation.
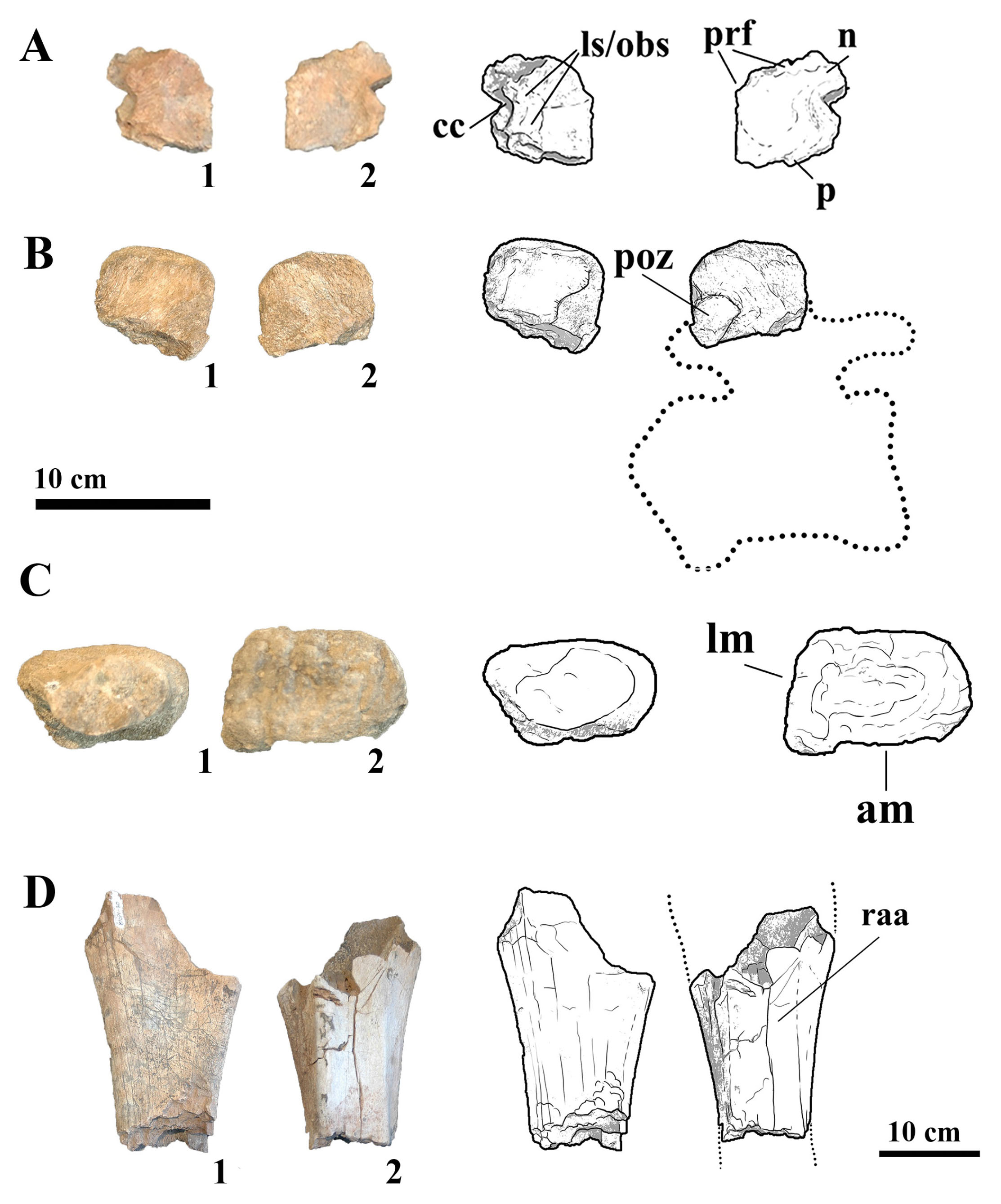
MMS-PV-78
This specimen comprises a fragmentary neural spine with both postzygapophyses preserved (Fig. 10B). The spine is elliptical in cross-section in anterior and ventral views and rectangular in lateral view. Although heavily weathered, the size and proportions of the spine are consistent with a middle caudal position. The elliptical cross-section is consistent with similar elements from Overosaurus paradasorum (Coria et al., 2013), although the overall spine is more vertically oriented than in Rinconsaurus caudamirus, which tends to exhibit more posteriorly inclined spines in the caudal region (Calvo et al., 2003).
MMS-PV-79
This specimen is an incomplete distal epiphysis of a probable metacarpal IV (Fig. 10C). In distal view, the articular surface is rectangular and rugose, with a slightly concave dorsal surface and a flattened ventral face. The distal facet suggests limited mediolateral mobility, consistent with weight-bearing functionality. This rectangular morphology is most comparable to Epachthosaurus sciuttoi (Martínez et al., 2004), which exhibits elongate metacarpals with asymmetrical distal facets. In contrast, saltasaurines such as Neuquensaurus australis and Saltasaurus loricatus possess more squared or subequal distal metacarpal facets (Otero, 2010).
MMS-PV-80
This specimen represents the proximal diaphysis of an ulna (Fig. 10D). The preserved radial articulation is slightly concave and maintains a constant mediolateral width along its height. In proximal view, the shaft exhibits a triradiate cross-section, a configuration considered typical for neosauropods, particularly within Titanosauriformes (Wilson & Sereno, 1998). The ulna is nearly straight in its preserved portion. This morphology is consistent with that observed in Dreadnoughtus schrani (Lacovara et al., 2014), a basal titanosauriform, and is also present in more derived saltasaurines such as Neuquensaurus (Otero, 2010).
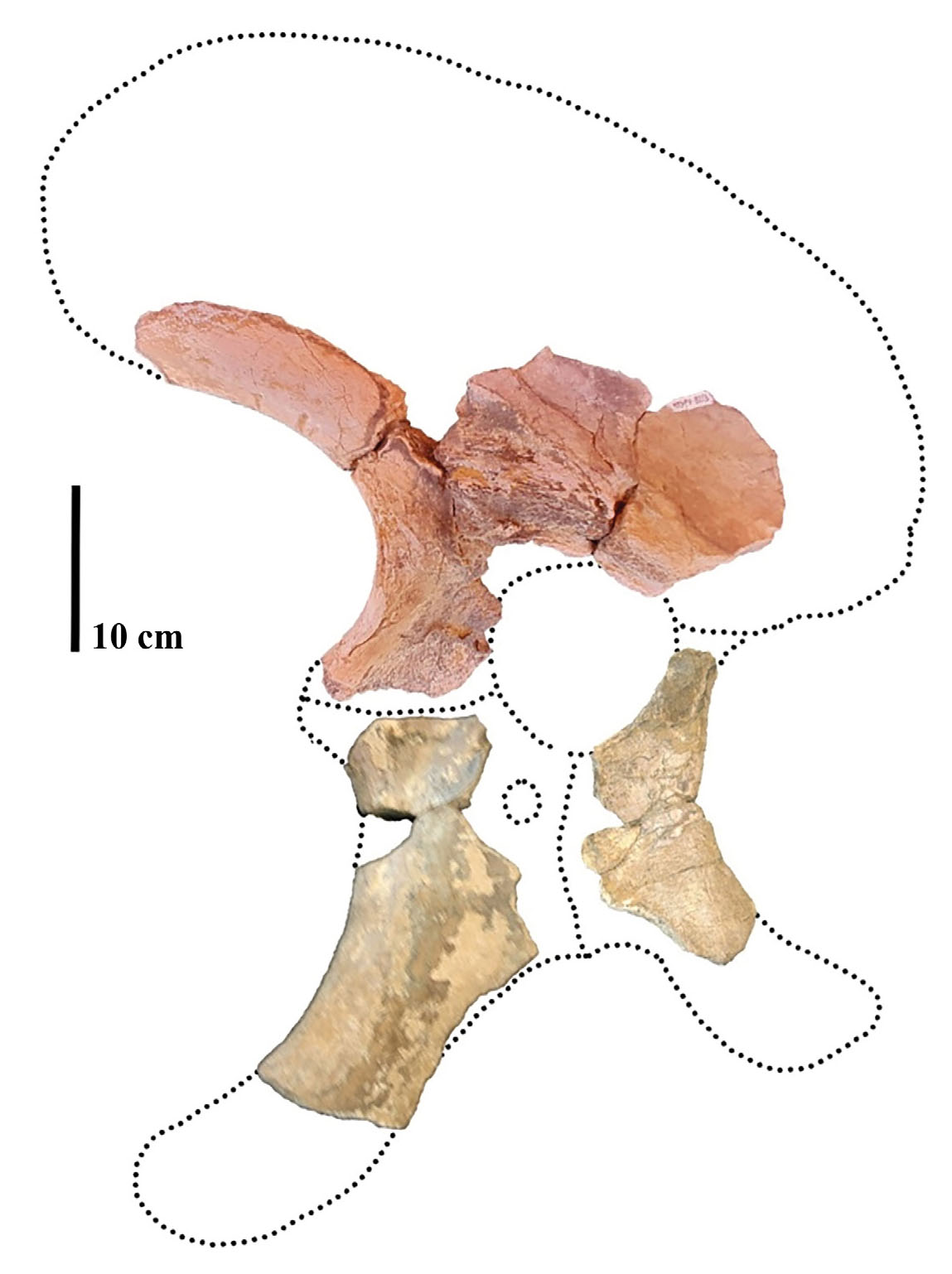
MMS-PV-82
This specimen includes three highly weathered and fragmented pelvic girdle elements, identified as a right ischium (MMS-PV-82/01), a right pubis (MMS-PV-82/02), and a left ilium (MMS-PV-82/03) (Fig. 11). The elements exhibit varying degrees of preservation due to differential exposure and diagenetic alteration. Although the majority of the external bone surface is eroded or obscured by cemented sedimentary matrix, several anatomical features permit meaningful taxonomic interpretation.
- Elements of the pelvic girdle of specimen MMS-PV-82 in suggested articulation (pubis and ischium are mirrored to reflect the ones from the same side of the Ilium).
MMS-PV-82/01: right ischium - The ischium is fragmented, preserving most of the acetabular margin and its iliac peduncle and lacking most of the distal portion of the shaft (Fig. 12A). The proximal portion, including the acetabular margin, is mediolaterally broader than the rest of the element. Notably, the iliac peduncle does not exhibit a constricted “neck” as in most titanosauriformes, is medially twisted like the one found in Futalongkosaurus dukei (Calvo et al., 2007) and is anteroposteriorly narrower than the distal shaft. This configuration resembles the ischial morphology of derived saltasaurids such as Saltasaurus loricatus (Bonaparte & Powell, 1980) and Neuquensaurus australis (Otero, 2010), in which the ischial shaft projects posteriorly at a nearly right angle to the acetabular region. No muscle scars or ischial tuberosities are visible, likely due to the weathered condition of the bone. Pneumatisation is absent, which is consistent with most known saltasaurines, in contrast to certain basal titanosaurs where pneumatic invasion of pelvic elements has been documented (Cerda et al., 2012).
- (A) Photographs and line drawings fragmented right Ischium MMS-PV-82/01 in (1) medial and (2) lateral views. Abbreviations: ilped, iliac peduncle; isped, ischiatic peduncle. (B) Photographs and line drawings of fragmented right pubis MMS-PV-82/02 in (1) medial and (2) lateral views. Abbreviations: acet, acetabulum; ilped, iliatic peduncle; isped, ischiatic peduncle. (C) Photographs and line drawings of fragmented left ilium MMS-PV-82/03 in (1) lateral and (2) medial views. Abbreviations: acet, acetabulum; ft, feeding traces; poap, postacetabular process; pped, pubic peduncle; prap, preacetabular process.
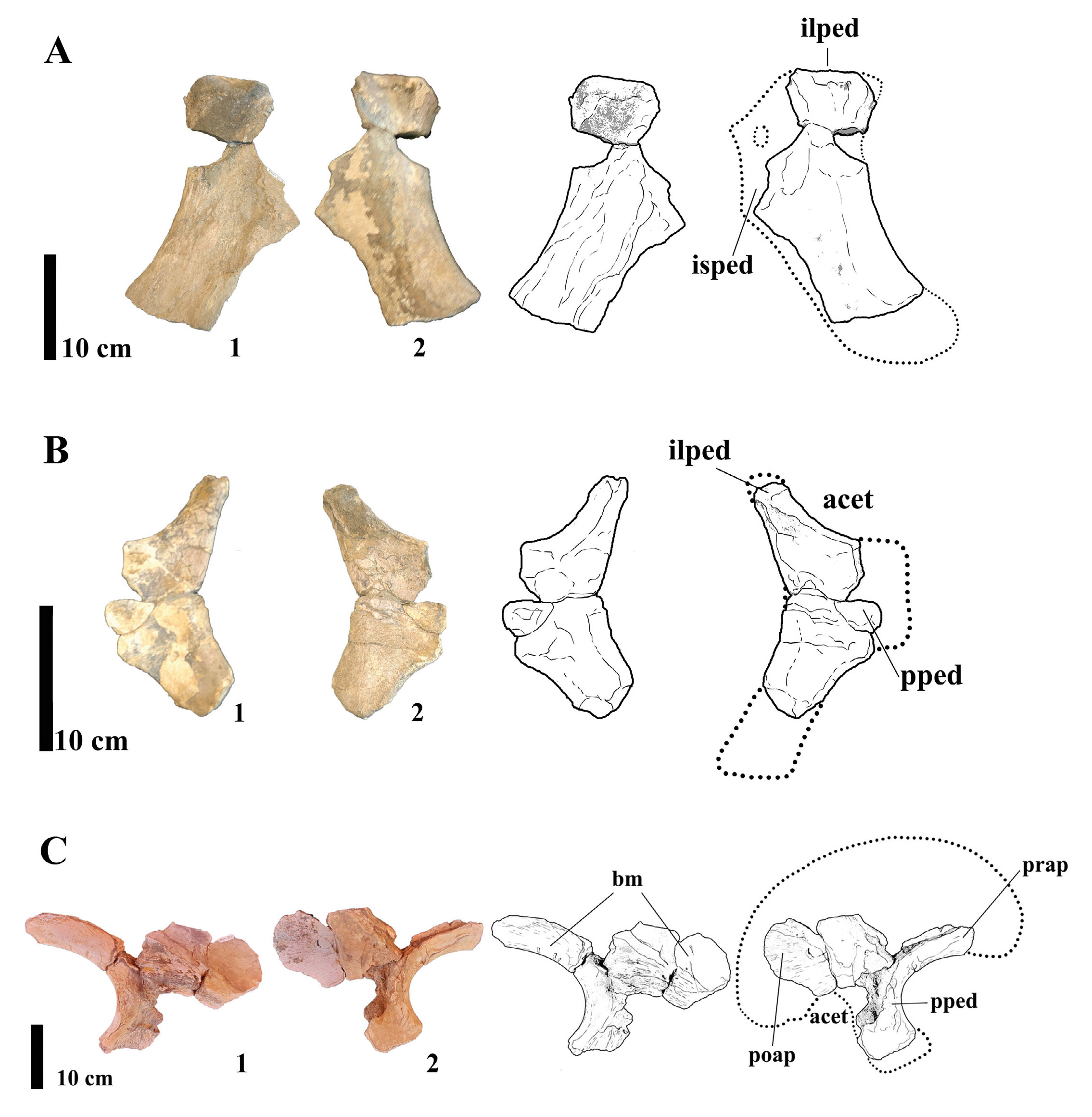
MMS-PV-82/02: right pubis - Only the proximal portion of the right pubis is preserved, including a partial iliac peduncle (Fig. 12B). This peduncle is dorsoventrally straight and features a distinct triangular depression just beneath it on the lateral surface. While this character is not diagnostic in isolation, it is morphologically similar to the proximal pubis of Dreadnoughtus schrani (Lacovara et al., 2014), where deep muscle attachment scars are also present proximally. The pubis lacks a longitudinal ridge, character found in Aeolosaurus, Opisthocoelicaudia and in Futalongkosaurus.
MMS-PV-82/03: left ilium - The left ilium is reconstructed from four fragments and includes the lower margin of the preacetabular process, part of the pubic process, and a section of the acetabular wall (Fig. 12C). The exposed internal structure doesn’t show any pneumatic features such as a camellate tissue. The preserved ventral margin of the preacetabular process is distinctly concave and aligned with the rest of the bone and curves downward toward its anterior tip, differently from the one of Overosaurus paradasorum, which is strongly deflected laterally along with the pubic peduncle. This morphology is consistent with saltasaurine titanosaurs, including Saltasaurus, Neuquensaurus, and Rocasaurus muniozi (Salgado & Azpilicueta, 2000), and has been proposed as a synapomorphy for Saltasaurinae (Gallina & Otero, 2009). The pubic process is dorsoventrally short, anteroposteriorly wide, and mediolaterally narrow. In distal view, the process has a D-shaped cross-section with a concave lateral and convex medial margin. This form is consistent with Dreadnoughtus schrani and Saltasaurus loricatus, differing from more rounded or robust morphologies in basal titanosaurian taxa such as Epachthosaurus sciuttoi (Martínez et al., 2004).
Feeding Traces on MMS-PV-82
Three discrete marks of potential biological origin are present on the ilium. These were assessed using the classification system of Hone & Watabe (2010), which categorises feeding traces based on morphology and inferred behaviour. The first mark is a shallow, drop-shaped groove on the lateral surface of the postacetabular process, nearly perpendicular to the long axis of the bone (Fig. 13A). This is interpreted as a score mark, resulting from tooth contact during soft tissue removal. The second mark consists of three deeper, transversely aligned punctures near the preacetabular process (Fig. 13B). These are classified as furrows, indicative of repetitive biting or scraping with greater force. The third mark, preserved on an undetermined associated fragment, is an oval depression interpreted as a pit (Fig. 13C), likely formed by localised pressure from a tooth tip without penetration.
- Feeding traces found on MMS-PV-82/03 interpreted as score marks (A), furrows (B) and a pit (C).

DISCUSSION
In this contribution we describe newly recovered titanosaurian material from the Bajo de la Carpa Formation at the Los Bastos locality (Neuquén Province, Argentina). The titanosaurian assemblage from the Los Bastos locality offers a valuable glimpse into the structure and evolutionary composition of sauropod faunas during the Santonian of northern Patagonia. The anatomical evidence reveals a dynamic and diverse sauropod community, with representatives morphologically compatible with both Colossosauria and Saltasauridae coexisting within the same depositional interval of the Bajo de la Carpa Formation. The specimen MMS-PV-75, an associated series of anterior to middle caudal vertebrae, closely resembles those of members of Aeolosaurinae, a derived clade within Colossosauria. This clade is defined by features including distoplatyan caudal articular surfaces, lateral bulging of the transverse processes in anterior caudals, and a dorsoventral flattening of the centra, traits all present in MMS-PV-75. Its morphology is consistent with that of Rinconsaurus caudamirus (Calvo et al., 2003) and Aeolosaurus rionegrinus (Powell, 2003), indicating that this group of slender-bodied, long-tailed titanosaurs had already radiated across southern Gondwana by the Santonian. The caudal sequence represented by MMS-PV-75 demonstrates a progressive transition in vertebral morphology from anterior to middle caudals, consistent with patterns observed in colossosaurian taxa. Notably, the procoelous-distoplatyan condition of the anterior vertebrae resembles that of Malarguesaurus florenciae (González Riga et al., 2008), a basal titanosauriform from the southern Neuquén Basin. In contrast, the more posterior elements of the new specimen’s caudal series show a shift toward stronger procoely, indicating morphological variation along the tail. Additional features, such as small pneumatopores and ventral bulges on the transverse processes, suggest affinities with Overosaurus paradasorum and Uberabatitan ribeiroi, all of which are considered members of the derived clade Aeolosaurinae (Filippi et al., 2019; Hechenleitner et al., 2020). The anterodorsal inclination of the anterior articular faces further aligns MMS-PV-75 with rinconsaurian and aeolosaurian taxa. Despite this distinctive combination of characters, the fragmentary nature of the material prevents the erection of a new taxon at this stage. Until more diagnostic elements are recovered and additional axial positions can be assessed, MMS-PV-75 is best referred to Titanosauria indet. within Colossosauria. The presence of such a derived aeolosaurine features supports the interpretation that Colossosauria underwent a significant diversification event before the Campanian (Filippi et al., 2019; Carballido et al., 2022). Similarly, MMS-PV-39 was attributed to Colossosauria, more specifically to Rinconsauria, which includes taxa such as Gondwanatitan faustoi and Trigonosaurus pricei. This placement is supported by the combination of an anteriorly placed neural arch, transversely convex ventral surface, and absence of pneumatic features, all characters that are congruent with those of basal colossosaurians. MMS-PV-76, although being an isolated element, points to a greater degree of titanosaurian morphological variability at the Los Bastos locality than previously recognised. Cranial material recovered from the locality, such as MMS-PV-77, supports the assignment of the specimen to Titanosauria, possibly within more derived clades. However, the incompleteness of the preserved elements precludes a more refined taxonomic identification at this time. A similar limitation applies to MMS-PV-78, a fragmentary neural spine interpreted as a middle caudal element of a titanosaurian sauropod. While its size and morphology are consistent with other colossosaurians, its fragmentary condition prevents further resolution. Additional postcranial remains, such as the partial metacarpal MMS-PV-79, lack diagnostic features sufficient for precise taxonomic identification but nonetheless support the presence of robust, non-saltasaurine titanosaurs within the Bajo de la Carpa palaeoecosystem. MMS-PV-82 shows features of the pelvic girdle consistent with taxa attributed to Saltasauridae, the most derived clade of Saltasauroidea, establishing MMS-PV-82 as the oldest saltasaurid currently known from the Neuquén Basin. The co-occurrence of colossosaurians and saltasaurids at Los Bastos indicates a degree of taxonomic overlap, with representatives of distinct titanosaurian lineages potentially occupying different ecological niches. This pattern echoes those observed in other Late Cretaceous ecosystems of South America, where taxonomic diversity among titanosaurs is paralleled by morphological divergence in traits related to locomotion, body size, and tail structure (Hechenleitner et al., 2020; Gallina et al., 2021). MMS-PV-75 was recovered in near-articulation, suggesting low-energy burial conditions and limited post-mortem transport, while MMS-PV-82 displays surface weathering and feeding traces attributable to theropods or crocodyliforms. The morphology, orientation, and distribution of these marks, comprising score marks, furrows, and a shallow pit, are consistent with theropod feeding behaviour, particularly that of abelisaurids or megaraptorids, both of which are known from the Bajo de la Carpa Formation (Filippi et al., 2016; Bellardini et al., 2021; Gianecchini et al., 2021; Porfiri et al., 2018), while not compatible with Unenlagids such as Diuqin (Porfiri et al., 2024) and other small theropods (e.g., Alvarezsaurians) from the same Formation. The pattern and spacing of the traces support their interpretation as the result of scavenging rather than active predation, especially in light of the absence of overlapping bite marks or extensive skeletal disarticulation (Fiorillo, 1991; Pobiner, 2008). While crocodyliforms are also present in the formation and capable of producing puncture and drag marks (Vasconcellos & Carvalho, 2010), the lack of drag scoring or crushing damage makes their involvement unlikely. These marks are consistent with scavenging behaviours previously documented in the overlaying Anacleto Formation (Bellardini et al., 2021), suggesting continuity in theropod–sauropod interactions across stratigraphic units. Taken together, the anatomical and preliminary taphonomic data from Los Bastos enhance our understanding of sauropod evolution in the Late Cretaceous of Gondwana. The newly described specimens not only broaden the known range of titanosaurian morphological variability but also reinforce the emerging view that titanosaur faunas were more taxonomically and ecologically diverse than previously recognised.
CONCLUSION
The here described titanosaurian fossils recovered from the Bajo de la Carpa Formation at the Los Bastos locality contribute to our understanding of sauropod diversity and evolution during the Santonian in northern Patagonia. Despite the fragmentary nature of the material, the detailed morphological descriptions and comparative analysis yielded valuable new data regarding the structure, variability, and evolutionary placement of mid-Cretaceous titanosaurs in Gondwana. The data from Los Bastos indicates overlapping diversification of multiple titanosaurian clades. Specimens MMS-PV-75 and MMS-PV-39 were both attributed to Colossosauria, with MMS-PV-75 showing strong affinities with Aeolosaurinae. Both specimens exhibit characteristic features such as procoelous-distoplatyan anterior caudals, transversely expanded transverse processes, and the absence of internal pneumatic structures. These features they share with the clade reinforce the hypothesis that colossosaurian diversity had already expanded across southern South America by the Santonian, with aeolosaurine morphology firmly established well-before the Campanian. In contrast, MMS-PV-82, comprising pelvic elements including an ischium, pubis, and ilium, was attributed to Saltasauridae, marking the earliest known occurrence of this clade in the Neuquén Basin. This extends the stratigraphic range of saltasaurids into the Santonian and could provide new insight into the early stages of their radiation. The feeding marks observed on MMS-PV-82 represent a useful ichnological evidence of trophic interactions, most likely involving theropods, enriching the behavioural and ecological framework of Late Cretaceous Patagonia. These data enhance our understanding of titanosaurian diversity, distribution, and palaeoecology and underscore the importance of underexplored localities and stratigraphic intervals in reconstructing the evolutionary history of South American dinosaurs.