

INTRODUCTION
The first Late Cretaceous dinosaur tracksite at Roztocze Hills in southeastern Poland found in 2005 near the village of Potok, in the outcrop exposed along the road from Stara Huta to Potok Sanderki. That site revealed hadrosaur trackway and the isolated theropod footprint (Gierliński et al., 2008). The second locality with various theropod and ornithischian track morphotypes was discovered in 2006 on the Młynarka Mountain (Fig. 1A; Gierliński, 2009, 2015). However, first vertebrate track described from Młynarka Mountain (known also as Szopowe quarry) was a Miocene perissodactyl print (Gierlinski et al., 2007) identified previously as cf. Palaeotheriipus, but later, in the opinion of Dalla Vecchia (personal communication, 2007), it represents rather a rhinocerotid footprint (i.e., Rhinoceripeda track). Dinosaur tracks occur few meters below the Miocene track-bearing horizon, at the bottom of the exposed section.
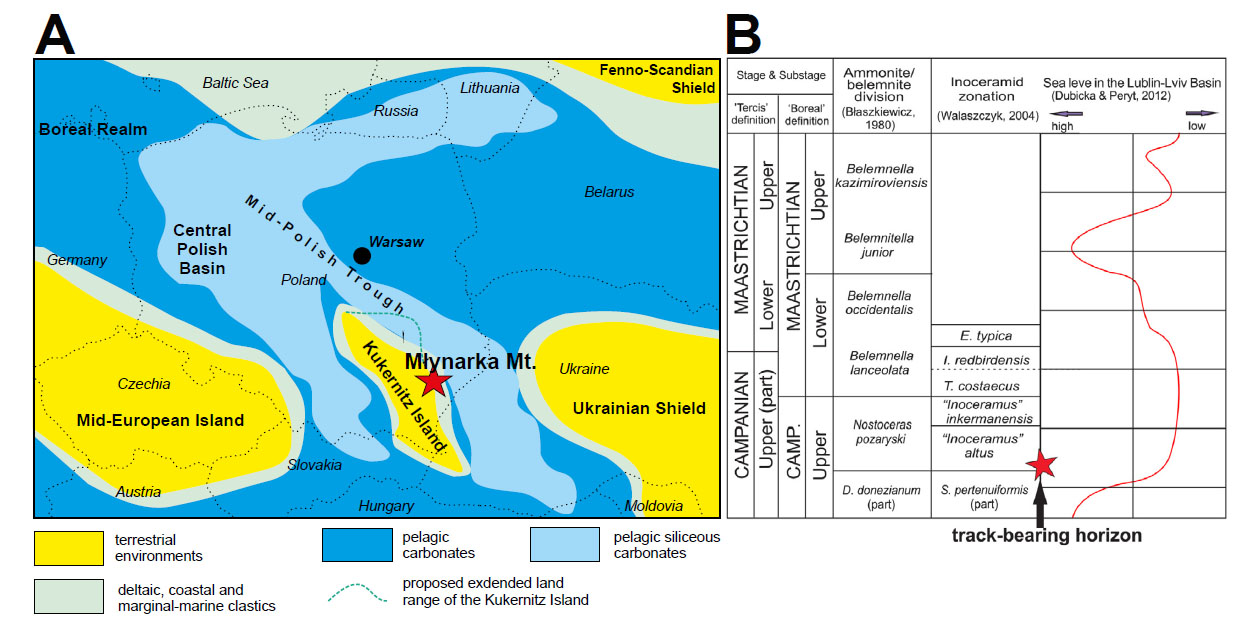
- Campanian palaeogeography of the central Europe (A) and stratigraphic position of the track-bearing horizon compared with sea-level curve of the Lublin-Lviv Basin (B), after Pasternat et al. (1987), Niechwedowicz et al. (2021) – modified and Gierliński (2015).
Moreover, the palaeogeography allows us to compare other vertebrate tracks found there during last two decades with tetrapod fauna recognized in surrounding regions, focusing especially on the Upper Cretaceous of northwestern Hungary, Haţeg and Transylvanian basins of Romania. We also made a revision of three track morphotypes formerly described from the Roztocze Hills.
GEOLOGICAL SETTING
During the Late Cretaceous, the area was part of the epicontinental Central Polish Basin. Palaeobiogeographically, it was located in the centre of the North European Province (Kauffman, 1973). The evolution of the Polish lowlands during this time was strongly influenced by eustatic changes and the inversion of the Mid-Polish Trough (Fig. 1A; Kauffman, 1973).
The subhercynian phase of Alpine orogenesis, from the latest Coniacian to Maastrichtian, caused the redevelopment of palaeotectonic-facies zones in southern Poland and southwestern Ukraine (Walaszczyk, 2003, 2012). Pasternak et al. (1987) indicated that this phase probably caused an uplift of the southeastern part of the Danish-Polish Trough. This uplifted emerged area was named Kukernitz Island by Pasternak et al. (1987). It was located in the area of the present-day Lower San Anticlinorium and the northern part of today’s eastern Carpathians and Carpathian Foredeep (Świdrowska et al., 2008). A global fall of the sea-level in the latest Campanian was indicated by several authors (Haq et al., 1988; Barrera and Savin, 1999; Li et al., 1999; Li et al., 2000; Friedrich et al., 2005; Ifrim & Stinnesbeck, 2010). The development of this land mass closed the Late Cretaceous basin in the Mid-Polish Trough from the south and southwest.
During the Late Cretaceous, the Roztocze Hills were located to the north of this uplifted area of Kukernitz Island and could be temporarily emerged (Świdrowska et al., 2008). In the Roztocze Hills, specifically in the quarry on Młynarka Mountain (Góra Młynarka; Fig. 1A), dinosaur footprints are preserved in arenaceous limestone Gierliński et al. (2008) and Gierliński (2009). These deposits represent sediments of the southeastern part of the Border Synclinorium, deposited in the Lviv-Stryj Basin. The Lviv-Stryj Basin was an epicontinental, carbonate platform basin. The age of the dinosaur track-bearing horizons at Młynarka Mt., initially referred to as Late Maastrichtian, was later reevaluated to the uppermost Campanian based on foraminifera taxa (Fig. 2B; Gierliński, 2015).
- Neoanomoepus isp., specimen JP Cr 740, from the Campanian of Młynarka Mountain, Poland (A) in comparison with the holotype of Neoanomoepus perigrinatus Lockley et al., 2009, specimen CU 199.22, from the Berriasian of British Columbia, Canada (B).

Analyses of palynofacies in the Middle Vistula River section, representing an epicontinental succession (Niechwedowicz et al., 2021), indicate predominantly neritic conditions, with a significantly offshore location suggested by a very low terrestrial/marine palynomorph ratio. However, the presence of terrestrial sporomorphs, freshwater and nearshore algae, and estuarine dinocysts, albeit in very low abundances, suggests proximity to Kukernitz Island.
The supply of terrestrially-sourced organic matter, likely from nearby Kukernitz Island, regularly increased water oxygenation and phytoplankton productivity in the marine basin. A short-term cyclicity in the palynofacies pattern, manifested as significant influxes of translucent phytoclasts (Tyson, 1993) probably reflects deltaic sedimentation that is assumed to have prevailed on the margins of Kukernitz Island during the late Campanian – early Maastrichtian (Niechwedowicz et al., 2021). Reworked dinocysts (Oxfordian to early Cenomanian) potentially redeposited from Kukernitz Island indicate that older strata were exposed and eroded on the island during this period (Niechwedowicz et al., 2021). The transport of material from the island could have been caused by changes in river runoff or even tectonic activity, which might have impacted Kukernitz Island and increased the basinward transport of terrestrial and nearshore material (Niechwedowicz et al., 2021).
MATERIAL AND METHODS
The new ichnite from the Młynarka Mountain presented here was found by the second author (SJ) in December 2024. The original specimen is so far left in situ at GPS coordinates: N 50°30’50.772”, E 23°3’42.78”. In the sites of Młynarka Mountain and Potok dinosaur tracks are preserved as the imprints (natural moulds). Normally the trackways are well preserved if the surrounding surface is not crashed. Any isolated tracks here presented are because of surface damage not because of taphonomy, nor natural weathering. In the Potok site, trampled surface is damaged by the road maintenance. At Młynarka Mountain, tracks are exposed on the surface constantly damage by motocross activity. And on other side, new tracks are constantly uncovered by construction activity of a neighbour ski station. The sites are occasionally patrolled by the Polish Geological Institute, Jurapark and Zagroda Guciów. Footprints are then photographed and casted using plaster or polymer clay (polyvinyl chloride). Two original footprints from Potok site are haused in Zagroda Guciów. Nine casts from Potok and Młynarka Mountain are stored in the collections of the Polish Geological Institute in Warsaw and Jurapark in Bałtów.
Photographs are taken strictly parallel to the surface by ultralow distortion Carl Zeiss lens, under controlled artificial low-key lighting diffused by an additional filter.
All measurements are based on standard methods (e.g., Haubold, 1971; Leonardi, 1987; Lallensack et al., 2025).
Abbreviations of cited repositories: CU-MWC - University of Colorado, Boulder, Colorado, USA; JP - JuraPark collection, Bałtów, Solec Kujawski and Krasiejów, Poland; LPB - Laboratory of Palaeontology of the Faculty of Geology and Geophysics, University of Bucharest, Bucharest, Romania; Muz. PIG - Gelogical Museum of the Polish Geological Institute-National Research Institute, Warsaw, Poland.
TRACK DESCRIPTION AND SYSTEMATIC AFFINITY
New Młynarka Mountain specimen JP Cr 740 (Fig. 2A) is the manus-pes set with clear heteropody. There is lack of distinct pad traces on the pes and manus. Functionally tetradactyl manus is slightly entaxonic and wider than long - 7 cm long and 11 cm wide. Digitigrade tridactyl pes with widely divaricated digits is as wide as long - 17 cm long and 17 cm wide. Manus is inwardly rotated and situated lateral to pes. Manus has short bulbous digit impressions. In contrast, the pedal digits are elongate, but relatively thick and elongated oval-shaped. Pedal digit II and III are equal in length – each one is 13 cm long. But digit IV is longer – 15 cm long. Angles between the pedal digits’ axes are as follow: II-III 39°, III-IV = 40 °. The pedal digits total divarication equals 79°. All three metatarsophalangeal pads are present and formed a bilobed “heel”. The footprint is then a very symmetrical in shape.
In the Cretaceous dinosaur tracks, the bilobed “heel” with the exception of Siamopodus Lockley et al., 2006 is typical for iguanodontid and hadrosaurid footprints (Currier et al., 1991; Lockley & Hunt, 1995). However, those large ornithopod footprints like Iguanodontipus Sarjeant et al., 1998, Caririchnium Leonardi, 1984 and Hadrosauropodus Lockley et al., 2003 are larger, most robust with broader digits and much smaller manus in comparison to the pes. Our specimen shows features similar to anomoepodid tracks (i.e., Anomoepodidae sensu Gierliński, 1991), such as highly divaricated pedal digits and subequal in length. Among anomoepodid tracks, Neoanomoepus Lockley et al., 2009 seems the best match. Despite the authors’ original description looking closely at the pedal ichnite of the manus-pes set no 2, in the holotype trackway A of Neoanomoepus perigrinatus Lockley et al., 2009 (specimen CU 199.22), we can see bilobed “heel” developed at the same discreet way like in our material from Młynarka Mountain (Fig. 2B). Moreover, in the cited manus-pes set no 2, in the specimen CU 199.22, the manus is tetradactyl, pes is tridactyl and that manus-pes association is nearly identical to the specimen JP Cr 740 from the Młynarka Mountain (see, Fig. 2). The specimen JP Cr 740 is only slightly larger with much wider divaricated pedal digits than CU 199.22. However, its measurements fit in between tracks parameters seen in the N. perigrinatus type material, where the total divarication of pedal digits varies between 35° and 81° and the pes length varies between 10 cm and 19 cm.
The tracks originally assigned to the ichnogenus Neoanomoepus came from Mist Mountain Formation (Berriasian in age) in Canada and Phra Wihan Formation (Berriasian-Valanginian) in Thailand. Additionally, Lockley et al. (2009) mentioned other Neoanomoepus-like material from Tithonian-Berriasian of Las Cerradicas in Spain and Dande Sandstone Formation (unknown age) in Zimbabwe. The latter one was more recently redescribed as Afrodelatorrichnus Bordy et al., 2020, supposed of heterodontosaurid origin (Gierliński, 2024).
McCrea et al. (2014) reported more tracks identical or similar to Neoanomoepus from the Minnes Group (Berriasiane-Valanginian) of Canada. And hitherto the youngest Neoanomoepus-like tracks are described as similar to Anomoepus by Henderson et al. (2022) from St. Mary River Formation (Campanian-Maastrichtian) of Canada.
Henderson et al. (2022) supposed thescelosaurid genus Parksosaurus known from neighbouring Horseshoe Canyon Formation as the possible trackmaker of those anomoepodid tracks from St. Mary River Formation. Following such coincidence correlation, the closest stratigraphically and geographically dinosaur faunas to search for the possible trackmaker of Młynarka Mt. anomoepodid track came from the Santonian of northwestern Hungary and from the Maastrichtian Haţeg and Transylvanian basins of Romania. Excluding Ceraratopsia accordingly to Czepiński & Madzia (2024), there are the members of Hadrosauridae, Rhabdodontidae, Nodosauridae, Titanosauria, Abelisauridae, Avialae, Oviraptosauria, Troodontidae, Dromaeosauridae and tetanurans (e.g., Benton et al., 2010; Botfalvai et al., 2015; Csiki-Sava et al., 2016).
Among the listed ornithopods, the rhabdodontids or rhabdodontomorphs in general rather than hadrosaurids might be considered as the possible trackmaker because of larger manus with less derived and more gracile pes.
The unnamed Early Cretaceous rhabdodontomorph from the Cameros Basin in Spain nick-named ‘Vegagete ornithopod’ and described by Dieudonné et al. (2016) is so far the unique specimen with nearly complete pedal bones. The ‘Vegagete ornithopod’ shows relatively narrow digits subequal in length (see Fig. 10 in Dieudonné et al., 2016) what fits discussed footprint from Młynarka Mt. as well as Neoanomoepus and anomoepodid pedal pattern in general (i.e., basal ornithischian foot pattern).
Alleged rhabdodontid tracks from the Maastrichtian of Transylvania (Romania) were reported by Vremir & Codrea (2002). However, their tracks’ drawings resemble “Amblydactylus” illustrated by Sternberg (1932) – seemingly large ornithopod track referred now to the large theropod ichnogenus Irenesauripus Sternberg, 1932 (see, Currie, 1983). Whereas, photos of this Romanian specimens resemble large theropod footprint (Fig. 3A) from the Potok site discussed in next chapter.
- Campanian dinosaur tracks from the Roztocze Hills in Poland discovered in 2005-2011: cf. Wakinyantanka isp. Muz. PIG 1704.II.2 (A) and Hadrosauropodus trackway Muz. PIG 1704.II.1 (B) from the Potok site; small Hadrosauropodus trackway JP Cr 741 (C), cf. Saurexallopus isp. Muz. PIG 1704.II.4 (D) and the part of Dromaeosauripus trackway Muz. PIG 1704.II.6 from the Młynarka Mountain (E).

ASSOCIATED TETRAPOD ICHNOTAXA
Large theropod footprint Muz. PIG 1704.II.2 from Potok site (Fig. 3A) was identified by Lockley et al. (2014) as similar to the erected by them ichnogenus Wakinyantanka from the Hell Creek Formation (Maastrichtian) of South Dakota and compared to very similar footprints found in the Hunter Canyon Formation (Campanian-Maastrichtian) of Colorado and the Blackhawk Formation (Campanian) of Utah, USA (see, Fig. 11 in Lockley et al., 2014). And they are also very similar to the tracks described from the Maastrichtian of Romania by Vremir & Codrea (2002). So, their trackmakers were probably some medium to moderately large tetanurans widely distributed in the Late Cretaceous world, or alternatively left by unrelative theropods with the same foot pattern due to convergence.
The Wakinyantanka-like track from Potok was associated by the trackway of large ornithopod (Fig. 3B) with pes 42 cm long and 47 cm wide This trackway was assigned to Hadrosauropodus by Gierliński et al. (2008). And the trackway with the footprints sharing the same morphology, but much smaller (pes is 17 cm long and 21 wide) came from Młynarka Mt. - specimen JP Cr 741 (Fig. 3C). Sometimes the same small and the same short-toed ornithopod tracks from the Lower Cretaceous of southeastern Asia were named as Ornithopodichnus by Kim et al. (2009). And then the same assignation was applied to the trackway from Młynarka Mt. by Gierliński (2015). However, short-toed Ornithopodichnus represents distinctive morphology among ornithopod tracks in the Lower Cretaceous, but not much different in comparison with the Late Cretaceous derived ornithopod footprints like Hadrosauropodus. Thus, in our recent opinion specimen JP Cr 741 is very similar to the Potok Hadrosauropodus, but just much smaller. It was possibly left by a juvenile or diminutive hadrosaurid.
The isolated uncatalogued ornithischian tetradactyl print from Młynarka Mt. supposed of ceratopsian affinity by Gierliński (2015), in the light of number of various ornithopods from the Late Cretaceous European archipelago might be the manus impression of one of them as well.
The Młynarka Mt. tracksite revealed also interesting theropod footprints. Gierliński (2008) reported track Muz. PIG 1704.II.4 similar to Saurexallopus Harris, 1997 (Fig. 3D) – a medium-sized bird-like track, possibly of oviraptorosaur origin as argued by Gierliński & Lockley (2013a).
Gierliński (2008) reported also from the same site the well-preserved dromaeosaur trackway (Fig. 3E) attributed previously to the ichnogenus Velociraptorichnus Zhen et al. (1994), what was later questioned by Lockley et al. (2016). Dromaeosaur tracks from Młynarka Mount indeed importantly differs from Velociraptorichnus by slightly larger size (about 16 cm in length), thicker digits and proximal portion of digit II not imprinted. Thus, our specimen is closer to Dromaeosauripus Kim et al. (2008), than to Velociraptorichnus.
The affinity of the next footprint (specimen Muz. PIG 1704.II.3 and JP Cr 664) is problematic. This large tetradactyl track (Fig. 4) was originally labelled as Macropodosaurus Zakharov,1964, by Gierliński (2008). First tracks of that kind were described by Zakharov (1964) from the Albian of Shirkent River valley in Tadzhikistan and later supposed of the therizinosaur origin by Sennikov (2006). Since 2008, Macropodosaurus, Macropodosaurus-like but not identified as Macropodosaurus and putative Macropodosaurus tracks were reported in the western North America, northern Africa and southeastern Asia. In 2013, Macropodosaurus (specimen CU 219.3) is described from the Turonian of Utah, USA (Gierliński & Lockley, 2013b). In 2017, track from Coniacian-Santonian of Morocco is assigned to Macropodosaurus (Masrour et al., 2017). In 2020, Macropodosaurus-like tracks from the Jinju Formation (possibly Aptian in age) of Korea were attributed to large bipedal crocodylomorphs and named Batrachopus grandis by Kim et al. (2020). A little later Sennikov (2021) showed that Batrachopus grandis and cf. Macropodosaurus, isp. from Utah are nearly identical and argued for their segnosaur (i.e., therizinosaur) origin. Moreover, Sennikov (2021) questioned assignment of B. grandis to Batrachopus because of being too different from Batrachopus genotype (i.e., B. deweyi Hitchcock, 1843).
- cf. Haenamichnus isp., the cast Muz. PIG 1704.II.3 and JP Cr 664 - probable azhdarchid footprint from the Campanian of Młynarka Mountain, Poland.

Having now a relatively rich material of tracks referred to Macropodosaurus we could see two discreetly different morphotypes among them. The footprint from Młynarka Mountain in Poland and that one from Morocco are paraxonic. They also show the space between digits and the bear-like shaped metapodium with subtriangular “heel”. Toes are relatively short and narrow parallelly arranged. In contrast, the type material from Tadzhikistan, the material from Utah and Korea comprises more theropod-like mesaxonic tracks with relatively long digits integrated with metapodium and subrectangular “heel”.
The footprints from Młynarka Mountain and Morocco rather resemble morphology exhibited by Haenamichnus gainensis Kim et al., 2012 - possible bipedal azhdarchid tracks from Haman Formation (Albian) of Korea. And even assuming the high degree of convergence between the footprints attributed to therizinosaurs and azhdarchid pterosaurs (Xing et al., 2025), the coincidence correlation speaks also in favour of azhdarchid affinity of the Młynarka Mt. specimen (Muz. PIG 1704.II.3). While there is a lack of therizinosaur remains in the Upper Cretaceous of Europe (Zanno, 2010), there is enough big pterosaur specimen LPB R.2347 from the Hațeg Basin (with predicted wingspan over 8 m) to produce such large, 35 cm in length footprint Muz. PIG 1704.II.3 (see, Vremir et al., 2018).
DISCUSSION
The morphology of newly discovered footprint matches ichnofamily Anomoepodidae Lull, 1953. Anomoepodids comprises commonly small and rarely medium bipedal and quadrupedal morphotypes characterized by the functionally tridactyl pes with the toes nearly equal in length, relatively short and widely divaricated. They generally reflect basal ornithischian foot pattern and show wide stratigraphic occurrence comprising the Late Triassic Eoanomoepus Lockley et al., 2018, the Jurassic Anomoepus Hitchcock, 1848 and the Cretaceous Neoanomoepus Lockley et al., 2009. This type of footprints was hitherto unknown in the Upper Cretaceous of Europe. In the Cretaceous of Europe, anomoepodid pedal morphology corresponds with the foot architecture seen among rhabdodontomorphs – the basal iguanodontian ornithopods. In the Late Cretaceous, the rhabdodontomorphs and particularly Rhabdodontidae Weishampel et al., 2003 were endemic to the Late Cretaceous European archipelago (Augustin et al., 2023). At that time, the studied area has been located at the northeastern coast of the Kukernitz Island - a part of this archipelago.
Tetrapod ichnotaxa from the Campanian of Roztocze Hills seems expectable for the Late Cretaceous European archipelago. Rhabdodontid, moderately large tetanuran, hadrosaurid, oviraptosaur, dromaeosaurid and azhdarchid tracks’ record corresponds with the fauna recognized in the neighbouring Haţeg and Transylvanian basins (e.g., Benton et al., 2010; Botfalvai et al., 2015; Csiki-Sava et al., 2016).
Foraminifers from the track-bearing horizon demonstrate that those sediments were deposited during the late Campanian. Thus, it is possible to suppose that during the time of a global lowstand, the studied region became temporary emerged. Naturally this view is strongly supported by finds of terrestrial vertebrate tracks in the Roztocze Hill. Moreover, the rich flora occurs in these sediments of the Roztocze region and the western Ukraine (Pasternak et al., 1987; Halamski, 2013). Complete compound leaves and abundance of angiosperm remains strongly lends no support for a long transport distance prior to burial and indicates nearby emergent vegetated areas (Halamski, 2013). This is also obvious that the Late Cretaceous huge tree-fern stem (88 cm high), being composed of numerous siphonostelic stems, could not have been transported far away in the sea (Gierliński et al., 2008).
Late Cretaceous epicontinental deposits in the Lviv-Stryi Basin (Moldovian, Moesian and Scythian platforms and Nort Dobrudjan orogen) was highly controlled by sea-level changes (Świdrowska et al., 2008). The patchy Late Cretaceous rock record of this area does not allow precise palaeogeographic and palaeotectonic reconstruction of this area. However, it is likely that many uplifted and denudated areas occurred there. For instance, during the Campanian time the shallow or emerged platform separated the Moldovian area in Romania and the Pre-Dobrogea Depression (Ukraine). The Central and South Dobrogea area during the Santonian-Maastrichtian was also uplifted and the sedimentation was mainly controlled by sea-level changes. The denudated lowland area occurred also during Coniacian and Santonian, between the Pre-Dobrogea Depression and the present Babadag Syncline, and in the eastern part of Moesian Plate. The uplifted edge of the East European Platform was located to the north-east of Lviv-Stryj Basin and its emergence was highly connected with sea-level changes. Since the latest Campanian, the global sea level is believed to be at the one of the lowest levels in the Late Cretaceous times. Thus, mentioned regions were emerged providing a dispersal routes from the East European Land to Kukernitz Island and to the peri-Tethys area.
CONCLUSIONS
The paper describes the first discovery of an anomoepodid track from the Upper Cretaceous of Europe. This track is assigned to the ichnogenus Neoanomoepus, previously known from the Lower Cretaceous of Canada, Thailand and potentially Spain. The Neoanomoepus footprints seem to be a track morphotype, which might be left by thescelosaurids as well as rhabdodontomorphs. And the coincidence correlation speaks in favour of its rhabdodontid affinity.
The paper also discusses associated tetrapod ichnotaxa found in the Roztocze Hills, including cf. Wakinyantanka isp., Hadrosauropodus isp., Dromaeosauripus isp. and cf. Saurexallopus. In addition, the large tetradactyl tetrapod footprint originally described as Macropodosaurus is compared to Haenamichnus and supposed of the giant pterosaur origin.
Our studies revealed the correspondences between the Roztocze (i.e., Kukernitz Island) tetrapod ichnoassemblage and as the Late Cretaceous tetrapod faunas of Romania, Hungary and other parts of the Late Cretaceous European archipelago, suggesting potential biogeographic connections. Furthermore, the occurene of terrestrial vertebrate traces extends the range of terrestrial conditions to the north-west, at least periodically during Campanian.